You might also like
- Isolation and Characterization of Ralstonia Solanacearum Causing Bacterial Wilt of Tomato in NigeriaDocument10 pagesIsolation and Characterization of Ralstonia Solanacearum Causing Bacterial Wilt of Tomato in NigeriaElvis GergesNo ratings yet
- Effect Aqueous Extract of Xanthium Strumarium L Andtrichoderma Viride Against Rhizctonia SolaniDocument6 pagesEffect Aqueous Extract of Xanthium Strumarium L Andtrichoderma Viride Against Rhizctonia SolaniTJPRC PublicationsNo ratings yet
- Abo-State Et Al., 2015Document7 pagesAbo-State Et Al., 2015Jordana KalineNo ratings yet
- 2012 Chandra JPPRDocument6 pages2012 Chandra JPPRknchandu1No ratings yet
- Agri - IJASR - Eco-Friendly Management of - Khalid Mohammed ShihabDocument10 pagesAgri - IJASR - Eco-Friendly Management of - Khalid Mohammed ShihabTJPRC PublicationsNo ratings yet
- Effect of Some Algal Filtrates and Chemical Inducers On Root-Rot Incidence of Faba BeanDocument7 pagesEffect of Some Algal Filtrates and Chemical Inducers On Root-Rot Incidence of Faba BeanJuniper PublishersNo ratings yet
- Biological Control of Sugarbeet Root Rot Caused by Sclerotium RolfsiiDocument4 pagesBiological Control of Sugarbeet Root Rot Caused by Sclerotium RolfsiinormanwillowNo ratings yet
- The Effect of Plant Extracts As Seed Treatments To Control Bacterial Leaf Spot of Tomato in TanzaniaDocument10 pagesThe Effect of Plant Extracts As Seed Treatments To Control Bacterial Leaf Spot of Tomato in TanzaniaAdita Nur FaoziyahNo ratings yet
- 30 36 PDFDocument7 pages30 36 PDFray m deraniaNo ratings yet
- Plant extracts inhibit white root disease fungusDocument8 pagesPlant extracts inhibit white root disease funguseifaNo ratings yet
- 10.1515 - ZNC 2009 5 618Document6 pages10.1515 - ZNC 2009 5 618Sona HaciliNo ratings yet
- Sciencedirect: Evaluating Effective Trichoderma Isolates For Biocontrol of RhizoctoniaDocument8 pagesSciencedirect: Evaluating Effective Trichoderma Isolates For Biocontrol of RhizoctoniaAde PutNo ratings yet
- Plant Pathogenicity Phenotyping of Ralstonia Solanacearum StrainsDocument17 pagesPlant Pathogenicity Phenotyping of Ralstonia Solanacearum StrainsAlberto MachoNo ratings yet
- PBB 4 079Document8 pagesPBB 4 079InnezNo ratings yet
- Molluscicidaleffectoftuba-Tuba (Jatrophacurcaslinn) Extracts On Golden Apple Snail (Pomaceacanaliculatalamarck)Document6 pagesMolluscicidaleffectoftuba-Tuba (Jatrophacurcaslinn) Extracts On Golden Apple Snail (Pomaceacanaliculatalamarck)DanaNo ratings yet
- Role of Phenols and Antioxidant Enzymes in Biocontrol of Fusarium Oxysporum Causing Fusarium Wilt of Arachis Hypogeae. L (Groundnut)Document10 pagesRole of Phenols and Antioxidant Enzymes in Biocontrol of Fusarium Oxysporum Causing Fusarium Wilt of Arachis Hypogeae. L (Groundnut)TJPRC PublicationsNo ratings yet
- White RootDocument8 pagesWhite RootMuhd Nur Fikri ZulkifliNo ratings yet
- G 1005024146Document6 pagesG 1005024146IOSRjournalNo ratings yet
- Mutagenic Effect of Sodium Azide On Seed Germination ofDocument7 pagesMutagenic Effect of Sodium Azide On Seed Germination offnazisNo ratings yet
- Some in Vitro Observations On The Biological Control of Sclerotium Rolfsii, A Serious Pathogen of Various Agricultural Crop PlantsDocument8 pagesSome in Vitro Observations On The Biological Control of Sclerotium Rolfsii, A Serious Pathogen of Various Agricultural Crop PlantsInternational Organization of Scientific Research (IOSR)No ratings yet
- 29 12.0215aDocument7 pages29 12.0215aYeiru Azael RatmareNo ratings yet
- tmpEAC2 TMPDocument11 pagestmpEAC2 TMPFrontiersNo ratings yet
- Nicotiana GlaucaDocument7 pagesNicotiana GlaucaNicolas Ignacio AllendeNo ratings yet
- Short note on the effects of seaweed extracts on fungal growthDocument3 pagesShort note on the effects of seaweed extracts on fungal growthmas'udNo ratings yet
- tmp9626 TMPDocument6 pagestmp9626 TMPFrontiersNo ratings yet
- Anees 2010Document11 pagesAnees 2010Amanda RafaelaNo ratings yet
- Pdis 2002 86 2 127Document4 pagesPdis 2002 86 2 127Aspin PianNo ratings yet
- Antimicrobial Activity of Trichoderma harzianumDocument8 pagesAntimicrobial Activity of Trichoderma harzianumEmaIžakovićNo ratings yet
- Isolation of Trichoderma Spp. and Determination of Their Antifungal, Biochemical and Physiological FeaturesDocument7 pagesIsolation of Trichoderma Spp. and Determination of Their Antifungal, Biochemical and Physiological Featurescrashoverride29187No ratings yet
- 14 Preeti Saini 64-68Document5 pages14 Preeti Saini 64-68Umesh MogleNo ratings yet
- Isolation and Characterization of Antibiotic Producing Streptomyces from SoilDocument7 pagesIsolation and Characterization of Antibiotic Producing Streptomyces from SoilMuhammad Umer FarooqNo ratings yet
- Antifungal Activity of Selected Plant Leaves Crude Extracts Against A Pepper Anthracnose Fungus, (Ascomycota: Phyllachorales)Document9 pagesAntifungal Activity of Selected Plant Leaves Crude Extracts Against A Pepper Anthracnose Fungus, (Ascomycota: Phyllachorales)MelanieNo ratings yet
- Antibacterial Activity of Lantana Camara Linn and Lantana: Montevidensis Brig Extracts From Cariri-Ceará, BrazilDocument3 pagesAntibacterial Activity of Lantana Camara Linn and Lantana: Montevidensis Brig Extracts From Cariri-Ceará, BrazilevilbioNo ratings yet
- (D2) Aib - 2019 - 0050Document9 pages(D2) Aib - 2019 - 0050Neder BlancoNo ratings yet
- Biological control of onion diseaseDocument7 pagesBiological control of onion diseaseJosé Matías ZapiolaNo ratings yet
- Antibacterial Activity of Drosera RotundifoliDocument3 pagesAntibacterial Activity of Drosera RotundifoliValGocinNo ratings yet
- 1 s2.0 S0362028X22084447 MainDocument9 pages1 s2.0 S0362028X22084447 MainTanti 01No ratings yet
- Colletotrichum Gloeosporioides From Mango Ataulfo: Morphological, Physiological, Genetic and Pathogenic AspectsDocument7 pagesColletotrichum Gloeosporioides From Mango Ataulfo: Morphological, Physiological, Genetic and Pathogenic AspectsresearchinbiologyNo ratings yet
- Abdi Et Al. 2008Document6 pagesAbdi Et Al. 2008argos1301No ratings yet
- Compatibility StudyDocument5 pagesCompatibility Studygem68megNo ratings yet
- Antagonistic Activity of Trichoderma Isolates Against Sclerotium Rolfsii: Screening of Efficient Isolates From Morocco Soils For Biological ControlDocument9 pagesAntagonistic Activity of Trichoderma Isolates Against Sclerotium Rolfsii: Screening of Efficient Isolates From Morocco Soils For Biological ControlSandro ManchegoNo ratings yet
- Plant and Soil 2000Document7 pagesPlant and Soil 2000Imam HafizhNo ratings yet
- Bacillus Thuringiensis (BT.) Against Meloidogyne Incognita: The Effect of Endotoxin Produced byDocument11 pagesBacillus Thuringiensis (BT.) Against Meloidogyne Incognita: The Effect of Endotoxin Produced byPythonnnnNo ratings yet
- Fakhrfeshani Et Al 2012Document7 pagesFakhrfeshani Et Al 2012argos1301No ratings yet
- Inhibitory and Killing Activities of Medicinal Plants Against Multiple Antibiotic-Resistant Helicobacter PyloriDocument8 pagesInhibitory and Killing Activities of Medicinal Plants Against Multiple Antibiotic-Resistant Helicobacter PyloriNyayu_FitrianiNo ratings yet
- Jurnal Herbisida 1Document17 pagesJurnal Herbisida 1IlallangGurunNo ratings yet
- Albuquerque 2014Document8 pagesAlbuquerque 2014Dayvid SilvaNo ratings yet
- 04 0380 Jagessar Antimicrobial Ns PDFDocument15 pages04 0380 Jagessar Antimicrobial Ns PDFJose Rodrigo Ardila BurgosNo ratings yet
- Applied Sciences: Ffect of Sulfadimethoxine, OxytetracyclineDocument15 pagesApplied Sciences: Ffect of Sulfadimethoxine, OxytetracyclineshradhaNo ratings yet
- CYJS (BS) - 15-44-91 (Phytotoxicity, Cytotoxicity and Antioxidant Activity A Inulifolium)Document9 pagesCYJS (BS) - 15-44-91 (Phytotoxicity, Cytotoxicity and Antioxidant Activity A Inulifolium)JuanManuelAmaroLuisNo ratings yet
- Bio-Rational Management of Whitefly (Bemisia Tabaci) For Suppressing Tomato Yellow Leaf Curl VirusDocument15 pagesBio-Rational Management of Whitefly (Bemisia Tabaci) For Suppressing Tomato Yellow Leaf Curl VirussarfarazNo ratings yet
- Urn Cambridge - Org Id Binary 20170213065555240-0185 S0028646X99003652 S0028646X9900365XaDocument14 pagesUrn Cambridge - Org Id Binary 20170213065555240-0185 S0028646X99003652 S0028646X9900365XaPini KomalasariNo ratings yet
- Bacteria From The Citrus Phylloplane Can Disrupt Cell-Cell Signalling in Xanthomonas Citri and Reduce Citrus Canker Disease SeverityDocument10 pagesBacteria From The Citrus Phylloplane Can Disrupt Cell-Cell Signalling in Xanthomonas Citri and Reduce Citrus Canker Disease Severityjuan carlos CaicedoNo ratings yet
- Applsci 09 02239Document12 pagesApplsci 09 02239Panagiotis EliopoulosNo ratings yet
- Crop Protectants 2015Document9 pagesCrop Protectants 2015joyeeta8No ratings yet
- Admin Vol36 1mDocument8 pagesAdmin Vol36 1mCivNo ratings yet
- Biochemical Defence Reactions of Black Pepper Varieties Against Colletotrichum Gloeosporioides Incitant of Anthracnose DiseaseDocument6 pagesBiochemical Defence Reactions of Black Pepper Varieties Against Colletotrichum Gloeosporioides Incitant of Anthracnose DiseaseShailendra RajanNo ratings yet
- Mass Production of AM Fungal Inoculum by Soil Based Pot CultureDocument5 pagesMass Production of AM Fungal Inoculum by Soil Based Pot CultureIJARBS JOURNALNo ratings yet
- 539 31-38 FinalDocument8 pages539 31-38 FinalDavidNo ratings yet
- 1771Document10 pages1771Mahmoud GaberNo ratings yet
- Nanotechnology 1Document30 pagesNanotechnology 1karthikkandaNo ratings yet
- 5 PDFDocument12 pages5 PDFMahmoud GaberNo ratings yet
- 14TOPSJDocument8 pages14TOPSJMahmoud GaberNo ratings yet
- Deguo Et AlDocument4 pagesDeguo Et AlBenjamin MabanagNo ratings yet
- Lysis ActivityDocument7 pagesLysis ActivityMahmoud GaberNo ratings yet
- Report 1Document114 pagesReport 1sirsinto100% (1)
- Petry, Nancy M - Behavioral Addictions - DSM-5 and Beyond-Oxford University Press (2016) PDFDocument257 pagesPetry, Nancy M - Behavioral Addictions - DSM-5 and Beyond-Oxford University Press (2016) PDFanang7No ratings yet
- PDF Dis StandardsDocument35 pagesPDF Dis StandardsEdén PastoraNo ratings yet
- Module Essential Newborn CareDocument26 pagesModule Essential Newborn CareMichtropolisNo ratings yet
- 1 s2.0 S0269749120365866 MainDocument10 pages1 s2.0 S0269749120365866 MainPEDRO NETO DOMINGOS SIGUIMNo ratings yet
- Avf Creation Minor Scrub 1Document4 pagesAvf Creation Minor Scrub 1Bianca SapugayNo ratings yet
- JIPMER 2000: Ans. B 2. Which of The Following Is Not A Content of The Pudendal CanalDocument48 pagesJIPMER 2000: Ans. B 2. Which of The Following Is Not A Content of The Pudendal CanalLakshmidevi.R.M R.MNo ratings yet
- BIOLOGY PP1 SERIES 2Document132 pagesBIOLOGY PP1 SERIES 2hashiimNo ratings yet
- Ampalaya Chips With Sweet DipDocument31 pagesAmpalaya Chips With Sweet DipJayson San Juan86% (7)
- Routine Medical Checkup PublicationDocument11 pagesRoutine Medical Checkup Publication백만호No ratings yet
- Stool Analysis GuideDocument28 pagesStool Analysis GuideRajeev PareekNo ratings yet
- 130248-Gc1 Immulite Global Menu Final-01971879Document2 pages130248-Gc1 Immulite Global Menu Final-01971879Jano GarciaNo ratings yet
- Benzene MSDSDocument12 pagesBenzene MSDSTom Jose KooduthottiyilNo ratings yet
- Cerebellar SyndromeDocument3 pagesCerebellar SyndromePirasan Traditional Medicine CenterNo ratings yet
- Gen Bio1 - Q1 - Module1 - WK1Document19 pagesGen Bio1 - Q1 - Module1 - WK1Teacher Charlyn VlogNo ratings yet
- Minor Scrub 3 PDFDocument3 pagesMinor Scrub 3 PDFMadel AcobNo ratings yet
- Yr. 1 Microbiology. LECDocument22 pagesYr. 1 Microbiology. LECrex367100% (1)
- CHAPTER 57 Anatomy and Surgical Access of The MediastinumDocument6 pagesCHAPTER 57 Anatomy and Surgical Access of The MediastinumAbeNo ratings yet
- Biology: Chapter 1 Invitation To BiologyDocument9 pagesBiology: Chapter 1 Invitation To Biologyamstrghtr94No ratings yet
- Aditya M Pattewar Et Al 2012Document7 pagesAditya M Pattewar Et Al 2012jsrajoyd_224488661No ratings yet
- Checklist 7 Step of Occupational Diagnosis - 2021-2022Document4 pagesChecklist 7 Step of Occupational Diagnosis - 2021-2022Putri NoviyantiNo ratings yet
- Asada 2Document33 pagesAsada 2Anonymous iNLNBVtpoNo ratings yet
- Thesis OphthalmologyDocument7 pagesThesis OphthalmologyJames Heller100% (2)
- Maharlika Highway, Brgy. Magsaysay, Lopez, Quezon: Lopez National Comprehensive High SchoolDocument5 pagesMaharlika Highway, Brgy. Magsaysay, Lopez, Quezon: Lopez National Comprehensive High SchoolJoenell CabungcalNo ratings yet
- Croup: Dr. Shveta Sethi Narula A.P. MicrobiologyDocument14 pagesCroup: Dr. Shveta Sethi Narula A.P. Microbiology12. Akshit AtwalNo ratings yet
- New Yorker MarchDocument86 pagesNew Yorker Marchloco100% (1)
- Top Communicable Diseases in The Philippines 2019Document4 pagesTop Communicable Diseases in The Philippines 2019Nicole cuencosNo ratings yet
- First Aid - CPR - AED Participant's ManualDocument196 pagesFirst Aid - CPR - AED Participant's Manualadamiam100% (2)
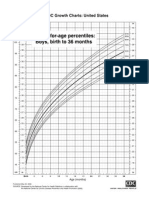
- CDC Boys 0 - 36 Mths - Length-For-AgeDocument1 pageCDC Boys 0 - 36 Mths - Length-For-AgepmverlaanNo ratings yet
- Surgical Abdomen of The Cattle - Descripcion Tecnica Laparotomia Flanco TernerosDocument32 pagesSurgical Abdomen of The Cattle - Descripcion Tecnica Laparotomia Flanco TernerosJhon Bustamante CanoNo ratings yet