You might also like
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (120)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Aero Ebook - Choosing The Design of Your Aircraft - Chris Heintz PDFDocument6 pagesAero Ebook - Choosing The Design of Your Aircraft - Chris Heintz PDFGana tp100% (1)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- Hormone QuestionnaireDocument8 pagesHormone QuestionnaireGabriel CamarenaNo ratings yet
- Hormone Cure Instructor Manual PDFDocument26 pagesHormone Cure Instructor Manual PDFGabriel Camarena83% (6)
- Sanskrit Subhashit CollectionDocument110 pagesSanskrit Subhashit Collectionavinash312590% (72)
- Cool Fire Manual 45M620N2UK 01 PDFDocument198 pagesCool Fire Manual 45M620N2UK 01 PDFPaun MihaiNo ratings yet
- SAFE RC Design ForDocument425 pagesSAFE RC Design ForMarlon Braggian Burgos FloresNo ratings yet
- Black Garlic ExtractDocument10 pagesBlack Garlic ExtractGabriel CamarenaNo ratings yet
- The Glucocorticoid Vulnerability HypothesisDocument20 pagesThe Glucocorticoid Vulnerability HypothesisGabriel CamarenaNo ratings yet
- Black Garlic - Alho NegroDocument9 pagesBlack Garlic - Alho NegroGustavo MartinsNo ratings yet
- C. Robert Mesle (Auth.) - John Hick's Theodicy - A Process Humanist Critique-Palgrave Macmillan UK (1991)Document168 pagesC. Robert Mesle (Auth.) - John Hick's Theodicy - A Process Humanist Critique-Palgrave Macmillan UK (1991)Nelson100% (3)
- UAV Design TrainingDocument17 pagesUAV Design TrainingPritam AshutoshNo ratings yet
- Case Study - Montana Mountain BikingDocument6 pagesCase Study - Montana Mountain Bikingbonny MishNo ratings yet
- The Diversity-Stability - McCann - NatureDocument6 pagesThe Diversity-Stability - McCann - Natureangstron328No ratings yet
- Diet Rapidly Alters The Human Gut MicrobiomeDocument26 pagesDiet Rapidly Alters The Human Gut MicrobiomeGabriel CamarenaNo ratings yet
- Climate Change Effects On ArbuscularDocument75 pagesClimate Change Effects On ArbuscularGabriel CamarenaNo ratings yet
- ElevatedatmosphericCO AltersrootsymbiontDocument12 pagesElevatedatmosphericCO AltersrootsymbiontGabriel CamarenaNo ratings yet
- Climate Change E¡ects On B Ene Cial Plant MicroorganisminteractionsDocument18 pagesClimate Change E¡ects On B Ene Cial Plant MicroorganisminteractionsGabriel CamarenaNo ratings yet
- ElevatedatmosphericCO AltersrootsymbiontDocument12 pagesElevatedatmosphericCO AltersrootsymbiontGabriel CamarenaNo ratings yet
- Aged Garlic ExtractDocument17 pagesAged Garlic ExtractGabriel CamarenaNo ratings yet
- Effects of Marijuana Smoking On PulmonaryDocument8 pagesEffects of Marijuana Smoking On PulmonaryGabriel CamarenaNo ratings yet
- Allium Thiosulfinates PDFDocument9 pagesAllium Thiosulfinates PDFGabriel CamarenaNo ratings yet
- Eating Behavior and StressDocument8 pagesEating Behavior and StressGabriel CamarenaNo ratings yet
- Nutrition Implications For Fetal Alcohol PDFDocument18 pagesNutrition Implications For Fetal Alcohol PDFGabriel CamarenaNo ratings yet
- Allium Thiosulfinates PDFDocument9 pagesAllium Thiosulfinates PDFGabriel CamarenaNo ratings yet
- Stress Food InflammationDocument5 pagesStress Food InflammationGabriel CamarenaNo ratings yet
- Oxidative Stress and Free Radicals in COPDDocument18 pagesOxidative Stress and Free Radicals in COPDGabriel CamarenaNo ratings yet
- Sensor Global de EstresDocument10 pagesSensor Global de EstresGabriel CamarenaNo ratings yet
- Subnutrition in The ElderlyDocument2 pagesSubnutrition in The ElderlyGabriel CamarenaNo ratings yet
- The Song of The NeanderthalDocument1 pageThe Song of The NeanderthalGabriel CamarenaNo ratings yet
- Open Access Definition of Inflammation, Causes of Inflammation PDFDocument9 pagesOpen Access Definition of Inflammation, Causes of Inflammation PDFGabriel CamarenaNo ratings yet
- Stress Exposure Food EmotionalDocument42 pagesStress Exposure Food EmotionalGabriel CamarenaNo ratings yet
- Depression: The Nutrition Connection: Invited PapersDocument8 pagesDepression: The Nutrition Connection: Invited PapersGabriel CamarenaNo ratings yet
- Depression: The Nutrition Connection: Invited PapersDocument8 pagesDepression: The Nutrition Connection: Invited PapersGabriel CamarenaNo ratings yet
- "Manganese-Induced Neurotoxicity PDFDocument20 pages"Manganese-Induced Neurotoxicity PDFGabriel CamarenaNo ratings yet
- 2.1. Inflammation and Pro-Inflammatory MediatorsDocument23 pages2.1. Inflammation and Pro-Inflammatory MediatorsGabriel CamarenaNo ratings yet
- Modification of Brain AgingDocument36 pagesModification of Brain AgingGabriel CamarenaNo ratings yet
- The First Voyage Round The World by MageDocument405 pagesThe First Voyage Round The World by MageGift Marieneth LopezNo ratings yet
- Chief Complaint: History TakingDocument9 pagesChief Complaint: History TakingMohamad ZulfikarNo ratings yet
- Introduction To HDLDocument28 pagesIntroduction To HDLBack UpNo ratings yet
- Multimedia System DesignDocument95 pagesMultimedia System DesignRishi Aeri100% (1)
- Lesson 1: Composition: Parts of An EggDocument22 pagesLesson 1: Composition: Parts of An Eggjohn michael pagalaNo ratings yet
- PGCRSM-01-BLOCK-03 Research Design ExperimentalDocument29 pagesPGCRSM-01-BLOCK-03 Research Design ExperimentalVijilan Parayil VijayanNo ratings yet
- Benefits and Limitations of BEPDocument2 pagesBenefits and Limitations of BEPAnishaAppuNo ratings yet
- Xtype Power Train DTC SummariesDocument53 pagesXtype Power Train DTC Summariescardude45750No ratings yet
- Community Architecture Concept PDFDocument11 pagesCommunity Architecture Concept PDFdeanNo ratings yet
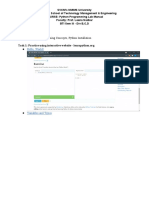
- Week - 2 Lab - 1 - Part I Lab Aim: Basic Programming Concepts, Python InstallationDocument13 pagesWeek - 2 Lab - 1 - Part I Lab Aim: Basic Programming Concepts, Python InstallationSahil Shah100% (1)
- Fortigate Firewall Version 4 OSDocument122 pagesFortigate Firewall Version 4 OSSam Mani Jacob DNo ratings yet
- 1500 Series: Pull Force Range: 10-12 Lbs (44-53 N) Hold Force Range: 19-28 Lbs (85-125 N)Document2 pages1500 Series: Pull Force Range: 10-12 Lbs (44-53 N) Hold Force Range: 19-28 Lbs (85-125 N)Mario FloresNo ratings yet
- Lancru hzj105 DieselDocument2 pagesLancru hzj105 DieselMuhammad MasdukiNo ratings yet
- Impact of Government Policies and EthicsDocument24 pagesImpact of Government Policies and EthicsGunveen AbrolNo ratings yet
- Possessive Determiners: A. 1. A) B) C) 2. A) B) C) 3. A) B) C) 4. A) B) C) 5. A) B) C) 6. A) B) C) 7. A) B) C)Document1 pagePossessive Determiners: A. 1. A) B) C) 2. A) B) C) 3. A) B) C) 4. A) B) C) 5. A) B) C) 6. A) B) C) 7. A) B) C)Manuela Marques100% (1)
- Liquitex Soft Body BookletDocument12 pagesLiquitex Soft Body Booklethello belloNo ratings yet
- Veritas™ High Availability Agent For WebSphere MQ Installation and Configuration Guide / WebSphere MQ InstallationDocument64 pagesVeritas™ High Availability Agent For WebSphere MQ Installation and Configuration Guide / WebSphere MQ InstallationkarthickmsitNo ratings yet
- CRISTIAN COLCERIU - PERSONALITATI CLUJENE Prof - Dr.ing - POMPILIU MANEADocument21 pagesCRISTIAN COLCERIU - PERSONALITATI CLUJENE Prof - Dr.ing - POMPILIU MANEAcristian colceriu100% (2)
- Nutrition 2022 PIRDocument22 pagesNutrition 2022 PIRAlmira LacasaNo ratings yet
- Transparency and Digitalization in The Public Administration of RomaniaDocument8 pagesTransparency and Digitalization in The Public Administration of RomaniaMădălina MarincaşNo ratings yet
- GLOBE2Document7 pagesGLOBE2mba departmentNo ratings yet
- Dynalift Sed0804679lDocument1 pageDynalift Sed0804679lzaryab khanNo ratings yet
- Sainik School Balachadi: Name-Class - Roll No - Subject - House - Assigned byDocument10 pagesSainik School Balachadi: Name-Class - Roll No - Subject - House - Assigned byPagalNo ratings yet