You might also like
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Jurnal Formulasi Rifampicin PDFDocument15 pagesJurnal Formulasi Rifampicin PDFKris Natalia ManihurukNo ratings yet
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- A C A D e M I C S C I e N C e SDocument6 pagesA C A D e M I C S C I e N C e SKris Natalia ManihurukNo ratings yet
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Formulation Development and Evaluation of Acyclovir Orally Disintegrating TabletsDocument5 pagesFormulation Development and Evaluation of Acyclovir Orally Disintegrating TabletsKris Natalia ManihurukNo ratings yet
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- ValganpkDocument13 pagesValganpkKris Natalia ManihurukNo ratings yet
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Mangostana L.) Pada Mencit Swiss (Muss Musculus) : Uji Efek Analgesik Ekstrak Kulit Manggis (GarciniaDocument7 pagesMangostana L.) Pada Mencit Swiss (Muss Musculus) : Uji Efek Analgesik Ekstrak Kulit Manggis (GarciniaKris Natalia ManihurukNo ratings yet
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- Jurnal Formulasi Rifampicin PDFDocument15 pagesJurnal Formulasi Rifampicin PDFKris Natalia ManihurukNo ratings yet
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Hesperidin and Hesperitin Preparation and Purification From Citrus Sinensis PeelsDocument4 pagesHesperidin and Hesperitin Preparation and Purification From Citrus Sinensis PeelsKris Natalia ManihurukNo ratings yet
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- 30 25 1 SMDocument10 pages30 25 1 SMMuhamad Wildan MalikNo ratings yet
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- Bio-Flavonoids With Promising Anti-Diabetic Potentials: A Critical SurveyDocument26 pagesBio-Flavonoids With Promising Anti-Diabetic Potentials: A Critical SurveyKris Natalia ManihurukNo ratings yet
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- Lecture 2 - ERTH 2403Document9 pagesLecture 2 - ERTH 2403Andre YimNo ratings yet
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Descendants of Joshua Seaver - 8 GenerationsDocument199 pagesDescendants of Joshua Seaver - 8 GenerationsRandy SeaverNo ratings yet
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Yearbook 2016Document308 pagesYearbook 2016achintNo ratings yet
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Water Quality ManagementDocument61 pagesWater Quality ManagementRahul RanjanNo ratings yet
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- Tutorial Nematoda 1 2019Document22 pagesTutorial Nematoda 1 2019Andre HimawanNo ratings yet
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- Primo Vessels Dr. AshrafDocument7 pagesPrimo Vessels Dr. AshrafAshraful IslamNo ratings yet
- Autonomic ReflexesDocument21 pagesAutonomic ReflexesJoe WeahNo ratings yet
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
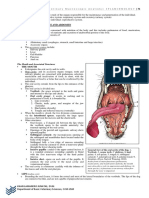
- Module 5 Anatomy I Splanchnology 1Document17 pagesModule 5 Anatomy I Splanchnology 1neannaNo ratings yet
- Pener1991, Locust Phase Polymorphism and Its EndocrineDocument79 pagesPener1991, Locust Phase Polymorphism and Its EndocrineAdriano Medeiros de SiqueiraNo ratings yet
- Experimental and Quasi-Experimental Designs For Generalized Causal Inference PDFDocument81 pagesExperimental and Quasi-Experimental Designs For Generalized Causal Inference PDFGonçalo SampaioNo ratings yet
- T Cell Development Thymic Education of T CellDocument34 pagesT Cell Development Thymic Education of T Cellapi-273068056No ratings yet
- 002MNSAlleles 1Document24 pages002MNSAlleles 1sherif abd el monemNo ratings yet
- Hongos Del ZamoranoDocument6 pagesHongos Del ZamoranoAnahí GallegosNo ratings yet
- 1,4 ButanediolDocument6 pages1,4 ButanediolVictor VikeneNo ratings yet
- 2019 Sec 1 Express Science SA2 East Spring SecondaryDocument26 pages2019 Sec 1 Express Science SA2 East Spring SecondaryMonali PNo ratings yet
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Bio 1M03Document73 pagesBio 1M03brady47No ratings yet
- Postdoc Cover Letter SampleDocument7 pagesPostdoc Cover Letter Samplevofysyv1z1v3100% (1)
- Kak UkkDocument4 pagesKak UkksantiNo ratings yet
- Biodiversity and Its ConservationDocument5 pagesBiodiversity and Its ConservationEenadu paperNo ratings yet
- Botany B.SCDocument2 pagesBotany B.SCAkash Deep PandeyNo ratings yet
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- Bio 2Document9 pagesBio 2triwicak15No ratings yet
- Water in The Human BodyDocument8 pagesWater in The Human BodyPaula AbadNo ratings yet
- Plant DescriptionDocument3 pagesPlant DescriptionJanine CamachoNo ratings yet
- Deav Chinhthuc Hsg12 Tinh-2021-2022Document15 pagesDeav Chinhthuc Hsg12 Tinh-2021-2022manhtuanktvn123No ratings yet
- A Survey of Color Charts For Biological DescriptionsDocument14 pagesA Survey of Color Charts For Biological DescriptionsgermporeNo ratings yet
- International Union For The Protection of New Varieties of PlantsDocument31 pagesInternational Union For The Protection of New Varieties of PlantsBimo HariokusumoNo ratings yet
- Final IRA - Mangoes From IndiaDocument174 pagesFinal IRA - Mangoes From Indiasakthi_dtsNo ratings yet
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Project Proposal: The Shinnecock Bay Restoration ProgramDocument17 pagesProject Proposal: The Shinnecock Bay Restoration ProgramDibyajyotiMandalNo ratings yet
- LipidDocument36 pagesLipidSina AgataNo ratings yet
- Jurnal Ilmiah Platax Vol. 6: (1), Januari 2018: Tongkaina and Bintauna Coastal Areas)Document12 pagesJurnal Ilmiah Platax Vol. 6: (1), Januari 2018: Tongkaina and Bintauna Coastal Areas)Shalltear BloodFallenNo ratings yet