You might also like
- 01 - Fisiología de La Hormona Antidiuretica (Reinhard)Document12 pages01 - Fisiología de La Hormona Antidiuretica (Reinhard)Rey Rodriguez67% (3)
- Constipacion FisiopatologiaDocument36 pagesConstipacion FisiopatologiaRoberto A. BermudezNo ratings yet
- Absorción con filtro BK como terapia eficaz en el riñón del mieloma múltipleFrom EverandAbsorción con filtro BK como terapia eficaz en el riñón del mieloma múltipleNo ratings yet
- Hidratación Parenteral en Pediatría PDFDocument6 pagesHidratación Parenteral en Pediatría PDFDavid OchoaNo ratings yet
- Bradiarritmias: Estación de SimulaciónDocument12 pagesBradiarritmias: Estación de SimulaciónOsnaider Andrés Cuello TorresNo ratings yet
- Causas y síntomas de la hipertensión endocraneanaDocument2 pagesCausas y síntomas de la hipertensión endocraneanaMariSol Pacheco MartinezNo ratings yet
- DESEQUILIBRIOS DEL SODIO Y CALCIODocument17 pagesDESEQUILIBRIOS DEL SODIO Y CALCIOLorena ZepedaNo ratings yet
- 2 2021 TRANSTORNOS ELECTROLITICOS DEL NA-K VamDocument38 pages2 2021 TRANSTORNOS ELECTROLITICOS DEL NA-K VamDaniela vocalNo ratings yet
- Hipertensión Portal para SeminarioDocument70 pagesHipertensión Portal para SeminarioCiro Calviño EstradaNo ratings yet
- Fisiologia Renal. Parte IIIDocument29 pagesFisiologia Renal. Parte IIIAlfonsoPetittoDeGregorioNo ratings yet
- Pediatria Cardio Tefy.Document34 pagesPediatria Cardio Tefy.Fabian Alexander Patino SalazarNo ratings yet
- Mieloma Multiple TratamientoDocument80 pagesMieloma Multiple TratamientoAlexis VegaNo ratings yet
- Alteraciones motilidad esofágicaDocument46 pagesAlteraciones motilidad esofágicaMakita CedeñoNo ratings yet
- Guias de Anticoagulación ChestDocument3 pagesGuias de Anticoagulación ChestCarlos AchurraNo ratings yet
- Exploracion Fisica Normal AdultoDocument2 pagesExploracion Fisica Normal AdultoDiego EspinozaNo ratings yet
- Encefalopatia HepáticaDocument16 pagesEncefalopatia HepáticaLaura C. SaltosNo ratings yet
- Vectors y Eje CardiacoDocument42 pagesVectors y Eje Cardiacojulio100% (1)
- HIPONATREMIADocument14 pagesHIPONATREMIAGustavo CarraroNo ratings yet
- Cap 62 Flujo Sanguineo CerebralDocument4 pagesCap 62 Flujo Sanguineo CerebralvictoramadoNo ratings yet
- Leucemias: clasificación, signos y tratamientoDocument8 pagesLeucemias: clasificación, signos y tratamientoCarlos Astete BórquezNo ratings yet
- Seminario-1 Edema CerebralDocument8 pagesSeminario-1 Edema CerebralANGELA ADRIANA RIVAS SALAZARNo ratings yet
- Manifestaciones Cutáneas de Linfomas y LeucimiasDocument66 pagesManifestaciones Cutáneas de Linfomas y LeucimiasinnovaciondocenteNo ratings yet
- Shock Cardiogenico LovesioDocument26 pagesShock Cardiogenico LovesioLaura HernandezNo ratings yet
- Manifestaciones Dermatologicas de Anemia FerropenicaDocument34 pagesManifestaciones Dermatologicas de Anemia FerropenicalecmcastilloNo ratings yet
- Encefalopatia HepaticaDocument4 pagesEncefalopatia HepaticaDavid PazNo ratings yet
- Crisis Hipertensiva y Hipertension en El Embarazo CardiologiaDocument27 pagesCrisis Hipertensiva y Hipertension en El Embarazo CardiologiaDaniela Emilia Javier SilfaNo ratings yet
- Sindrome Hiperglucemico Hiperosmolar No CetosicoDocument5 pagesSindrome Hiperglucemico Hiperosmolar No CetosicoHeiser LlanosNo ratings yet
- Neoplasia Endocrina MultipleDocument2 pagesNeoplasia Endocrina MultipleClaudia VargasNo ratings yet
- Insuficiencia Renal AgudaDocument41 pagesInsuficiencia Renal AgudaEstefanía P. HerreraNo ratings yet
- Síndrome coronario agudo: causas, síntomas y tratamientoDocument44 pagesSíndrome coronario agudo: causas, síntomas y tratamientoMarco Antonio Salazar PalaciosNo ratings yet
- Ecv-Medicina InternaDocument55 pagesEcv-Medicina InternaEngel MatamorosNo ratings yet
- Caso Clinico Neumonia IntrahosspitalariaDocument13 pagesCaso Clinico Neumonia IntrahosspitalariaBrayanEduardoTenorioYnfantesNo ratings yet
- Choque 4Document32 pagesChoque 4Alvaro BasilioNo ratings yet
- Retinopati Üa Diabe Ütica y EmbarazoDocument15 pagesRetinopati Üa Diabe Ütica y EmbarazoInternational Medical PublisherNo ratings yet
- MEHU130 - U2 - M5 - T3 - Glomerulopatías Primarias y Secundarias PDFDocument37 pagesMEHU130 - U2 - M5 - T3 - Glomerulopatías Primarias y Secundarias PDFHarold Flores MendozaNo ratings yet
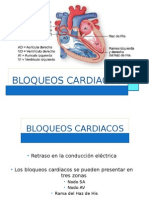
- Bloqueos cardiacos: tipos y características en EKGDocument17 pagesBloqueos cardiacos: tipos y características en EKGMariana ArmentaNo ratings yet
- Sindrome Coranario AgudoDocument42 pagesSindrome Coranario Agudopedro escorzaNo ratings yet
- Bloqueo de RamaDocument6 pagesBloqueo de Ramamigue5000No ratings yet
- Enfermedad Renal Crónica APSDocument17 pagesEnfermedad Renal Crónica APSAndrésVergaraJacksonNo ratings yet
- StrokeDocument110 pagesStrokeRodolfo Reséndiz100% (1)
- Seminario Shock 2018 InternosDocument53 pagesSeminario Shock 2018 InternosKelvin RonnielNo ratings yet
- NEUMOTÓRAXDocument13 pagesNEUMOTÓRAXCinthia Ordoñez0% (1)
- Corazón PDF 1Document145 pagesCorazón PDF 1Omar Leonardo Guerrero SuarezNo ratings yet
- Taquiarritmias 3Document23 pagesTaquiarritmias 3Sofía PinzónNo ratings yet
- Anemia MEGALOBLASTICADocument30 pagesAnemia MEGALOBLASTICAjoseNo ratings yet
- Cirugia No Cardiaca en Paciente CardiacoDocument19 pagesCirugia No Cardiaca en Paciente CardiacoGustavo Xavier Ayarza PulidoNo ratings yet
- Hemorragia Digestiva Alta y BajaDocument38 pagesHemorragia Digestiva Alta y Bajarodpin19No ratings yet
- Inmunología Del Trasplante. Vías de Rechazo (Presentación)Document10 pagesInmunología Del Trasplante. Vías de Rechazo (Presentación)Monikeit Tardígradus100% (1)
- Hepatitis Aguda Por FarmacosDocument26 pagesHepatitis Aguda Por FarmacosJosmaira FontainesNo ratings yet
- Infarto Agudo Al Miocardio STDocument5 pagesInfarto Agudo Al Miocardio STapi-3699557100% (5)
- Insuficiencia Cardiaca UrgenciasDocument49 pagesInsuficiencia Cardiaca UrgenciasJose angel Rojas martinez100% (1)
- ANTIARRITMICOSDocument6 pagesANTIARRITMICOSKaren LeónNo ratings yet
- Diureticos-Isaza Ocr PDFDocument10 pagesDiureticos-Isaza Ocr PDFSantiago Figueroa TorresNo ratings yet
- SIADH causa más común de hiponatremiaDocument14 pagesSIADH causa más común de hiponatremiaEsteban Montoya-VeraNo ratings yet
- Exantemas Pediatria PDFDocument66 pagesExantemas Pediatria PDFCarol ParraNo ratings yet
- Trastornos motores esofágicosDocument5 pagesTrastornos motores esofágicosPablo BosaNo ratings yet
- El Envenenamiento Por Mordedura en Centroamerica 2016Document34 pagesEl Envenenamiento Por Mordedura en Centroamerica 2016Álvaro RuedaNo ratings yet
- 13hernias de La Pared AbdominalDocument34 pages13hernias de La Pared AbdominalÁlvaro RuedaNo ratings yet
- La Entalpia de Descomposición Del Peróxido de Hidrogeno Lab 5 de FQ PDFDocument3 pagesLa Entalpia de Descomposición Del Peróxido de Hidrogeno Lab 5 de FQ PDFÁlvaro Rueda100% (1)
- Aislamiento Del GlucógenoDocument7 pagesAislamiento Del GlucógenoÁlvaro Rueda0% (1)
- 12APENDICITISDocument32 pages12APENDICITISÁlvaro RuedaNo ratings yet
- Punto Isoelectrico de Aminoácidos y ProteínasDocument8 pagesPunto Isoelectrico de Aminoácidos y ProteínasÁlvaro RuedaNo ratings yet
- TAREA 1 - HISTORIA NATURAL y MEDIDAS PREVENTIVASDocument1 pageTAREA 1 - HISTORIA NATURAL y MEDIDAS PREVENTIVASÁlvaro RuedaNo ratings yet
- Efectos de La Anemia en El Embarazo-1 PDFDocument7 pagesEfectos de La Anemia en El Embarazo-1 PDFÁlvaro RuedaNo ratings yet
- Análisis Cualitativo de LípidosDocument11 pagesAnálisis Cualitativo de LípidosÁlvaro RuedaNo ratings yet
- Hematuria IntramedDocument6 pagesHematuria Intramedtatis_naNo ratings yet
- Analisis de Sales Biliares y Ensayo Enzimatico para La Determinacion Del Colesterol SanguineoDocument18 pagesAnalisis de Sales Biliares y Ensayo Enzimatico para La Determinacion Del Colesterol SanguineoÁlvaro RuedaNo ratings yet
- Analisis de La Orina PDFDocument9 pagesAnalisis de La Orina PDFÁlvaro RuedaNo ratings yet
- 1-Efectos de Los Contaminantes Biologicos de Los AlimentosDocument3 pages1-Efectos de Los Contaminantes Biologicos de Los AlimentosÁlvaro RuedaNo ratings yet
- Expo Colestero BioquimicaDocument30 pagesExpo Colestero BioquimicaÁlvaro RuedaNo ratings yet
- EdemaDocument4 pagesEdemaÁlvaro RuedaNo ratings yet
- EdemaDocument4 pagesEdemaÁlvaro RuedaNo ratings yet
- EdemaDocument4 pagesEdemaÁlvaro RuedaNo ratings yet
- Nicolas CopérnicoDocument3 pagesNicolas CopérnicoÁlvaro RuedaNo ratings yet
- Meza Esparta Julia MargaritaDocument187 pagesMeza Esparta Julia MargaritaYelsin CamargoNo ratings yet
- Inyectores DieselDocument12 pagesInyectores DieselAlvaro Alexander Aliaga AvilaNo ratings yet
- Articulo de Hamburguesa, TerminadoDocument8 pagesArticulo de Hamburguesa, TerminadoAdix Adelia BernillaNo ratings yet
- Monografia APC Del TabacoDocument8 pagesMonografia APC Del TabacoNabil SutarNo ratings yet
- Orden de Entrada o InspeccionDocument1 pageOrden de Entrada o InspeccionEstefanía de FagundezNo ratings yet
- Teorías Psicológicas Del DueloDocument2 pagesTeorías Psicológicas Del Duelodon fer92% (12)
- Pauta Triptico Literario.Document2 pagesPauta Triptico Literario.daniela camposNo ratings yet
- Modulo de Fisica Grado 10Document35 pagesModulo de Fisica Grado 10YRDZ TUTOSNo ratings yet
- Guia Armarios TEKPANDocument14 pagesGuia Armarios TEKPANOmarNo ratings yet
- Cuantías de ObraDocument3 pagesCuantías de ObraRosa Casal CasapNo ratings yet
- Correlación de Pearson y regresión linealDocument16 pagesCorrelación de Pearson y regresión linealAlexander GarciaNo ratings yet
- TOPCART2016 L5 Observacion v20170222Document252 pagesTOPCART2016 L5 Observacion v20170222juanhk69100% (1)
- Metodo de Explotacion Corte y Relleno AscendenteDocument30 pagesMetodo de Explotacion Corte y Relleno Ascendenteabelkrusnik02100% (1)
- Registró Auxiliar 6 EdaDocument6 pagesRegistró Auxiliar 6 EdaWalter Quispe AymaNo ratings yet
- La Aceptacion de Si Mismo - Romano Guardini PDFDocument9 pagesLa Aceptacion de Si Mismo - Romano Guardini PDFmindyNo ratings yet
- 19 - Service Twister - Encendido - WWW - Clubtwister.com - ArDocument11 pages19 - Service Twister - Encendido - WWW - Clubtwister.com - ArlagorkNo ratings yet
- Tarea 5Document8 pagesTarea 5ROSA CLEVEDONo ratings yet
- Silabo - COMUNICACIÓN IIDocument8 pagesSilabo - COMUNICACIÓN IINerilzon JairNo ratings yet
- Comprención y Redacción Grupo 8Document6 pagesComprención y Redacción Grupo 8Marisol CarpioNo ratings yet
- Experimento de Luis PasteurDocument3 pagesExperimento de Luis PasteurdianaNo ratings yet
- Derivadas Logarítmicas y TrigonometricasDocument6 pagesDerivadas Logarítmicas y TrigonometricasMichelle MarinNo ratings yet
- Guía PrácticaElSecretoLeydeAtraccionDocument15 pagesGuía PrácticaElSecretoLeydeAtraccionAlberto MarpezNo ratings yet
- Comprimidos Obtenidos Por Compresión de Un GranuladoDocument12 pagesComprimidos Obtenidos Por Compresión de Un GranuladoMaria Rene TorresNo ratings yet
- Porcelagres antideslizanteDocument2 pagesPorcelagres antideslizanteJuliana RodriguezNo ratings yet
- Tecnicas para El Desarrollo de La CreatividadDocument16 pagesTecnicas para El Desarrollo de La CreatividadFrancisca JavierNo ratings yet
- Mercadona AnalisisDocument13 pagesMercadona Analisisicastrou50% (2)
- Aspectos Generales de Un PIPDocument13 pagesAspectos Generales de Un PIPPedro J. Zorrilla CaveroNo ratings yet
- Glosario Teoría Del Derecho PDFDocument8 pagesGlosario Teoría Del Derecho PDFLuis Mariano Betrian Aldea100% (1)
- IMPRESIONES EN PX EDENTULODocument38 pagesIMPRESIONES EN PX EDENTULOPau Santi100% (2)
- 200 Tareas Terapia BreveDocument5 pages200 Tareas Terapia BrevePremarthi PrabhuNo ratings yet