You might also like
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- No. Type of The Shell Deffect Description: Gross CracksDocument1 pageNo. Type of The Shell Deffect Description: Gross CracksekosaputrobbppbatuNo ratings yet
- Tafsir Ibnu Katsir Surat Al 'AbasaDocument9 pagesTafsir Ibnu Katsir Surat Al 'AbasaIwan Rahdianto100% (1)
- Tugas TaqiDocument3 pagesTugas TaqiekosaputrobbppbatuNo ratings yet
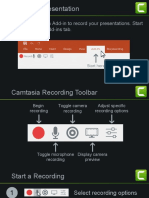
- Camtasia Getting Started Guide (Autosaved)Document4 pagesCamtasia Getting Started Guide (Autosaved)ekosaputrobbppbatuNo ratings yet
- Desain GoatDocument1 pageDesain GoatekosaputrobbppbatuNo ratings yet
- ReferencesDocument3 pagesReferencesekosaputrobbppbatuNo ratings yet
- Ante PostDocument22 pagesAnte PostekosaputrobbppbatuNo ratings yet
- Alokasi Kamar Pesonna MalioboroDocument4 pagesAlokasi Kamar Pesonna MalioboroekosaputrobbppbatuNo ratings yet
- CXP 057eDocument33 pagesCXP 057eArunee KongpengNo ratings yet
- Pola Label No. 112Document1 pagePola Label No. 112ekosaputrobbppbatuNo ratings yet
- Tabulasi Data Lapang Telur-ReniDocument27 pagesTabulasi Data Lapang Telur-ReniekosaputrobbppbatuNo ratings yet
- Kuesioner Telur EnglishDocument6 pagesKuesioner Telur EnglishekosaputrobbppbatuNo ratings yet
- Egg WashingDocument7 pagesEgg WashingekosaputrobbppbatuNo ratings yet
- Land and property financial recordsDocument2 pagesLand and property financial recordsekosaputrobbppbatuNo ratings yet
- Artificial Insemination (AI) of CattleDocument2 pagesArtificial Insemination (AI) of Cattleekosaputrobbppbatu100% (1)
- Albert EinsteinDocument2 pagesAlbert EinsteinekosaputrobbppbatuNo ratings yet
- Prosedur Mixed ModelDocument1 pageProsedur Mixed ModelekosaputrobbppbatuNo ratings yet
- Proses EkstruksiDocument10 pagesProses EkstruksiekosaputrobbppbatuNo ratings yet
- Dosen FPP UNDIPDocument10 pagesDosen FPP UNDIPekosaputrobbppbatuNo ratings yet
- Tek DGG Lanjut S-2#2Document11 pagesTek DGG Lanjut S-2#2ekosaputrobbppbatuNo ratings yet
- Teknologi Daging Lanjut Advanced Technology of Meat: Master Course in Animal Science v. Priyo BintoroDocument18 pagesTeknologi Daging Lanjut Advanced Technology of Meat: Master Course in Animal Science v. Priyo BintoroekosaputrobbppbatuNo ratings yet
- Perpustakaan BSNDocument1 pagePerpustakaan BSNekosaputrobbppbatuNo ratings yet
- Curing Smoking S-2#3Document12 pagesCuring Smoking S-2#3ekosaputrobbppbatuNo ratings yet
- Perubahan SFT Olahan Daging 1#S2Document13 pagesPerubahan SFT Olahan Daging 1#S2ekosaputrobbppbatuNo ratings yet
- Curing Smoking S-2#3Document12 pagesCuring Smoking S-2#3ekosaputrobbppbatuNo ratings yet
- Daftar PustakaDocument13 pagesDaftar PustakaekosaputrobbppbatuNo ratings yet
- Meat Products #4Document11 pagesMeat Products #4ekosaputrobbppbatuNo ratings yet
- Bacterial Starter Cultures For Meat FermentationDocument8 pagesBacterial Starter Cultures For Meat FermentationekosaputrobbppbatuNo ratings yet
- Antioxidant Effects of Broccoli Powder Extract in Goat Meat NuggetsDocument6 pagesAntioxidant Effects of Broccoli Powder Extract in Goat Meat NuggetsekosaputrobbppbatuNo ratings yet
- Reducing Sodium Levels in Frankfurters Using A Natural Flavor EnhancerDocument10 pagesReducing Sodium Levels in Frankfurters Using A Natural Flavor EnhancerekosaputrobbppbatuNo ratings yet
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (894)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- PMLSP 2 ReviewerDocument38 pagesPMLSP 2 ReviewerSophia Mae ClavecillaNo ratings yet
- Daftar PustakaDocument3 pagesDaftar PustakaAdnanNo ratings yet
- Antinfek 10HDocument20 pagesAntinfek 10Hmistytewest100% (1)
- Pemphigus Subtypes Clinical Features Diagnosis andDocument23 pagesPemphigus Subtypes Clinical Features Diagnosis andAnonymous bdFllrgorzNo ratings yet
- Cosmeceutic ALS: Cosmetics and PharmaceuticalsDocument19 pagesCosmeceutic ALS: Cosmetics and PharmaceuticalssowjanyaNo ratings yet
- Pecutan Akhir Science 2021Document29 pagesPecutan Akhir Science 2021Azween SabtuNo ratings yet
- Chiro Guidelines WHODocument51 pagesChiro Guidelines WHOMinh MinhNo ratings yet
- DNA TimelineDocument2 pagesDNA TimelineMinaMilićNo ratings yet
- Citric Acid Production Stirred TankDocument9 pagesCitric Acid Production Stirred TankKarliiux MedinaNo ratings yet
- Amazing Facts - Exercises 1Document3 pagesAmazing Facts - Exercises 1Alina DorobatNo ratings yet
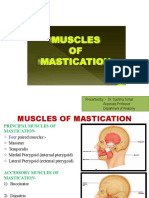
- Presented By:-Dr. Sushma Tomar Associate Professor Department of AnatomyDocument20 pagesPresented By:-Dr. Sushma Tomar Associate Professor Department of AnatomyCristine EchaveNo ratings yet
- Starship Troopers Skinnies Army BookDocument66 pagesStarship Troopers Skinnies Army BookDougBirtles100% (5)
- Lesson 7 - Organ TransplantationDocument14 pagesLesson 7 - Organ TransplantationJosee100% (1)
- Biology of Implant OsseointegrationDocument11 pagesBiology of Implant OsseointegrationAmIrNo ratings yet
- Effects of Climate on Living Things"This title "TITLE "Effects of Climate on Living ThingsDocument2 pagesEffects of Climate on Living Things"This title "TITLE "Effects of Climate on Living ThingsMelannie Magalong100% (1)
- Man Refined Energetic Seduction - Chris BaleDocument63 pagesMan Refined Energetic Seduction - Chris BaleSanti Sica0% (2)
- LWT - Food Science and TechnologyDocument8 pagesLWT - Food Science and TechnologyAzb 711No ratings yet
- 8 Cell - The Unit of Life-NotesDocument6 pages8 Cell - The Unit of Life-NotesBhavanya RavichandrenNo ratings yet
- Agilent ASMS 2019 ThP762 PosterDocument4 pagesAgilent ASMS 2019 ThP762 PostersppNo ratings yet
- Thailand Orchids Producer & Exporter Kultana Orchids Product ListDocument45 pagesThailand Orchids Producer & Exporter Kultana Orchids Product ListMalakh Raziel0% (2)
- Biomedical Physics, Vol 7 @medphyslibDocument360 pagesBiomedical Physics, Vol 7 @medphyslibStats -MANNo ratings yet
- Behaviour of Great Apes Studied by Three Female ScientistsDocument12 pagesBehaviour of Great Apes Studied by Three Female ScientistsSani AhmadNo ratings yet
- 1.4 - Diffusion and Osmosis - CrosswordPuzzleDocument1 page1.4 - Diffusion and Osmosis - CrosswordPuzzlemmssthNo ratings yet
- This Topic. So Congratulations Because Today I'm Going To Talk About TheDocument3 pagesThis Topic. So Congratulations Because Today I'm Going To Talk About TheJoon Bok NamleeNo ratings yet
- Lab Exercise 0ne: Carbohydrate Analysis Lab A.1 (Page 28)Document31 pagesLab Exercise 0ne: Carbohydrate Analysis Lab A.1 (Page 28)Goh Kae HorngNo ratings yet
- 01 MDCAT SOS Regular Session (5th June-2023) With LR..Document5 pages01 MDCAT SOS Regular Session (5th June-2023) With LR..bakhtawarsrkNo ratings yet
- Everyday Sports Injuries The Essential Step-By-Step Guide To Prevention, Diagnosis, and TreatmentDocument274 pagesEveryday Sports Injuries The Essential Step-By-Step Guide To Prevention, Diagnosis, and TreatmentAnonymous f6goFflg3T100% (10)
- Book of Mythical BeastsDocument32 pagesBook of Mythical BeastsRishabhNo ratings yet
- Full Download Human Physiology From Cells To Systems 8th Edition Lauralee Sherwood Solutions ManualDocument36 pagesFull Download Human Physiology From Cells To Systems 8th Edition Lauralee Sherwood Solutions Manualgambolrapinous.ggqcdr100% (39)
- Human Contact May Reduce Stress in Shelter DogsDocument5 pagesHuman Contact May Reduce Stress in Shelter DogsFlorina AnichitoaeNo ratings yet