You might also like
- Clase B1Document24 pagesClase B1andreaNo ratings yet
- Clase A2 y 3Document53 pagesClase A2 y 3andreaNo ratings yet
- Rotafolio Educativo para El Puerperio y La Crianza Temprana CHCC y PSM 3 PDFDocument42 pagesRotafolio Educativo para El Puerperio y La Crianza Temprana CHCC y PSM 3 PDFDanny Martinez RosalesNo ratings yet
- Anestésicos Locales en Odontología 2019Document44 pagesAnestésicos Locales en Odontología 2019andreaNo ratings yet
- Farmacologia DermatologicaDocument33 pagesFarmacologia Dermatologicaandrea100% (3)
- Vías de Administración y FF 2019Document48 pagesVías de Administración y FF 2019andreaNo ratings yet
- Apuntes (2) Topica. Curso Consejo Andalucia 2Document72 pagesApuntes (2) Topica. Curso Consejo Andalucia 2Javier Rodriguez MolinaNo ratings yet
- Clase A1Document40 pagesClase A1andreaNo ratings yet
- AngelesenlatierraDocument50 pagesAngelesenlatierraandreaNo ratings yet
- CLASE 001 - Patologia Cardiovascular Parte 1Document49 pagesCLASE 001 - Patologia Cardiovascular Parte 1andreaNo ratings yet
- Farmaco PediatriaDocument7 pagesFarmaco PediatriaEddopp100% (1)
- Farmacologia Antibioticos-Antifungicos - Antivirales (Clase 15)Document229 pagesFarmacologia Antibioticos-Antifungicos - Antivirales (Clase 15)claulskNo ratings yet
- 8.1) Preparacion de FleboclisisDocument20 pages8.1) Preparacion de Fleboclisisandrea100% (1)
- Anestésicos Locales en Odontología 2019Document44 pagesAnestésicos Locales en Odontología 2019andreaNo ratings yet
- 5.1) Farmacos AntiinflamatoriosDocument30 pages5.1) Farmacos AntiinflamatoriosandreaNo ratings yet
- 3.1) Farmacologia SNCDocument28 pages3.1) Farmacologia SNCandreaNo ratings yet
- Toma MuestraDocument42 pagesToma MuestraandreaNo ratings yet
- 3.4) Farmacologia EndocrinaDocument35 pages3.4) Farmacologia EndocrinaandreaNo ratings yet
- Norma Tecnica para La Atencion Integral en El Puerperio 2015Document297 pagesNorma Tecnica para La Atencion Integral en El Puerperio 2015pamela mendez100% (1)
- 5 09 Abril - 1 - El Cuidado - 1Document6 pages5 09 Abril - 1 - El Cuidado - 1andreaNo ratings yet
- Manual Obstetricia Ginecologia v5 2014Document661 pagesManual Obstetricia Ginecologia v5 2014Constanza100% (2)
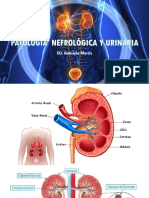
- Patologia Nefrologica y UrinariaDocument47 pagesPatologia Nefrologica y UrinariaandreaNo ratings yet
- Clase 2 Generalidades IaasDocument72 pagesClase 2 Generalidades IaasandreaNo ratings yet
- 8 Trabajo de Parto PrematuroDocument21 pages8 Trabajo de Parto PrematuroandreaNo ratings yet
- Niveles de intervención familiar en saludDocument28 pagesNiveles de intervención familiar en saludandreaNo ratings yet
- 4estrategias de AtenciónDocument9 pages4estrategias de AtenciónandreaNo ratings yet
- Sindrome Hipertensivo Del Embarazo (SHE) : CFT Santo Tomás - TensDocument28 pagesSindrome Hipertensivo Del Embarazo (SHE) : CFT Santo Tomás - Tensandrea100% (1)
- Asepsia y Antisepsia P - 2Document39 pagesAsepsia y Antisepsia P - 2andreaNo ratings yet
- Generalidades Del Proceso de EsterilizaciónDocument56 pagesGeneralidades Del Proceso de Esterilizaciónandrea100% (2)
- Apunte Salud Mental: Juan Videla A. Prof - Biología-Enfermero Magíster en EducaciónDocument8 pagesApunte Salud Mental: Juan Videla A. Prof - Biología-Enfermero Magíster en EducaciónandreaNo ratings yet
- Proteinas 11111Document19 pagesProteinas 11111Antonio M ChavezNo ratings yet
- Guía de Estudio. Sistema Digestivo - 1° ParteDocument5 pagesGuía de Estudio. Sistema Digestivo - 1° Parteroxana livaNo ratings yet
- Estructura y clasificación de proteínasDocument4 pagesEstructura y clasificación de proteínasSantiago Rosso JiménezNo ratings yet
- Banqueo BiologiaDocument94 pagesBanqueo Biologiabriyithe del pilar flores julcaNo ratings yet
- Que Es El Tranporte ActivioDocument8 pagesQue Es El Tranporte ActivioYennys Daza SalinasNo ratings yet
- Biología Celular Membrana Celular I 2 16Document48 pagesBiología Celular Membrana Celular I 2 16yurfaNo ratings yet
- Transporte de MacromoléculasDocument5 pagesTransporte de MacromoléculasFabi RodríguezNo ratings yet
- Aminoacidos Peptidos y ProteinasDocument27 pagesAminoacidos Peptidos y ProteinasjulyNo ratings yet
- Guia 5Document9 pagesGuia 5carlos contrerasNo ratings yet
- INFORME N°3: PLANOS CRISTALOGRAFICOS / Laboratorio de CristalografiaDocument26 pagesINFORME N°3: PLANOS CRISTALOGRAFICOS / Laboratorio de CristalografiaRonald Richard Quiliche GalvezNo ratings yet
- Química Biológica Clase 4 ProteínasDocument67 pagesQuímica Biológica Clase 4 ProteínasSergio ForgioneNo ratings yet
- Cuestionario Biologia ListoDocument4 pagesCuestionario Biologia ListodenissvillalobosrNo ratings yet
- Membrana CelularDocument1 pageMembrana CelularAmidNo ratings yet
- Resumen Fisiologia Solemne 1Document109 pagesResumen Fisiologia Solemne 1Maria PazNo ratings yet
- Estructura Cuaternaria de Las ProteínasDocument2 pagesEstructura Cuaternaria de Las Proteínaswimperlopez514No ratings yet
- PRIMER PARCIAL MODELO - RespuestasDocument9 pagesPRIMER PARCIAL MODELO - RespuestasDelfina RappaNo ratings yet
- Microdominios de MembranaDocument2 pagesMicrodominios de Membranakatykath1No ratings yet
- Tema 3Document6 pagesTema 3Chris melanie BazanNo ratings yet
- Apuntes Biologia SuerteDocument3 pagesApuntes Biologia SuerteCarolina Alexandra ZEBALLOS LIPANo ratings yet
- Menbrana CelularDocument6 pagesMenbrana CelularValery FreittesNo ratings yet
- Taller Transporte A Través de Membrana CelularDocument1 pageTaller Transporte A Través de Membrana CelularLiliana HernándezNo ratings yet
- Capítulo 3 - Trastornos Del Sistema InmuneDocument42 pagesCapítulo 3 - Trastornos Del Sistema InmuneConstanza ZentenoNo ratings yet
- Resumen Del Aparato DigestivoDocument2 pagesResumen Del Aparato DigestivoARMY BTSNo ratings yet
- 0 FacbfDocument2 pages0 FacbfSara CastroNo ratings yet
- Cristalografia MineralDocument45 pagesCristalografia MineralEddie Rodriguez BarraNo ratings yet
- Guía de membrana celularDocument8 pagesGuía de membrana celularNuria CabreraNo ratings yet
- Fisicoquímica de medicamentos y receptores celularesDocument9 pagesFisicoquímica de medicamentos y receptores celularesBrenda Alessandra Villanueva ChavezNo ratings yet
- Inmunología: preguntas sobre componentes del sistema inmunitario y respuesta inflamatoriaDocument11 pagesInmunología: preguntas sobre componentes del sistema inmunitario y respuesta inflamatoriavirginiaNo ratings yet
- 1er PARCIAL FISIOLOGÍA 2019Document5 pages1er PARCIAL FISIOLOGÍA 2019Marcelo GutierrezNo ratings yet
- Unidad 2 - Mecanismos de Respuestas A Nivel CelularDocument4 pagesUnidad 2 - Mecanismos de Respuestas A Nivel CelularNoelia OsunaNo ratings yet