You might also like
- Guía Trabajos Prácticos de BIOQUÍMICA 2021Document22 pagesGuía Trabajos Prácticos de BIOQUÍMICA 2021Emy CativaNo ratings yet
- CromatinaDocument15 pagesCromatinaSerch de la PazNo ratings yet
- Teorico 3 Primera Semana de DesarrolloDocument14 pagesTeorico 3 Primera Semana de DesarrolloSamantha RusinqueNo ratings yet
- Fases Del Ciclo CelularDocument8 pagesFases Del Ciclo CelularJazmin Alejandra GaiborNo ratings yet
- Ciclo Celular+Document18 pagesCiclo Celular+Yoel GutierrezNo ratings yet
- Tema. Citología. Ciclo y Divisiones CelularesDocument9 pagesTema. Citología. Ciclo y Divisiones Celularesmgonate4814No ratings yet
- Ciclo CelularDocument6 pagesCiclo Celularwert addNo ratings yet
- Anexo Ciclo CelularDocument5 pagesAnexo Ciclo Celularjuan lopezNo ratings yet
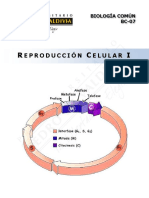
- BC07-15 Reproducción Celular I-WEBDocument19 pagesBC07-15 Reproducción Celular I-WEBCony SJNo ratings yet
- Ciclo celular, mitosis y meiosisDocument12 pagesCiclo celular, mitosis y meiosisVictor CrNo ratings yet
- CICLO CELULARrrrrDocument10 pagesCICLO CELULARrrrrEliana Itati BenitezNo ratings yet
- EMBRIOLOGIA ResumenDocument39 pagesEMBRIOLOGIA ResumenFran Madriaza50% (4)
- La HistogenesisDocument14 pagesLa HistogenesisCory Carver0% (1)
- Division CelularDocument11 pagesDivision CelularDiana ReinalesNo ratings yet
- Unidad II Resumen de EmbrioDocument7 pagesUnidad II Resumen de EmbrioAldana CuevasNo ratings yet
- Tarea 2 - Angeline Hurel Laina 2-5 HDocument12 pagesTarea 2 - Angeline Hurel Laina 2-5 HAngeline HurelNo ratings yet
- Histoembriologia Del Sistema AuditivoDocument18 pagesHistoembriologia Del Sistema Auditivofelipe paredes panichineNo ratings yet
- Fisiologia MitosisDocument5 pagesFisiologia MitosisErick Patiño MuñozNo ratings yet
- Desarrollo embrionario tempranoDocument8 pagesDesarrollo embrionario tempranoAndrés PachónNo ratings yet
- CelulaDocument4 pagesCelulaGenesis LizethNo ratings yet
- CicloCelularDocument12 pagesCicloCelularYiber Milena Olarte CNo ratings yet
- La Ressignificación Del Gemelo SobrevivienteDocument53 pagesLa Ressignificación Del Gemelo Sobrevivientepaola garcia100% (1)
- Desarrollo Embrionario.Document10 pagesDesarrollo Embrionario.salomendNo ratings yet
- GastrulacionDocument2 pagesGastrulacionJannine Jiménez de GonzálezNo ratings yet
- Clase de Transporte Intracelular UDA 2020Document3 pagesClase de Transporte Intracelular UDA 2020Celina RiveraNo ratings yet
- Division CelularDocument5 pagesDivision CelularElvira SaraviaNo ratings yet
- Embriología: desarrollo prenatalDocument163 pagesEmbriología: desarrollo prenataldavidNo ratings yet
- Ciclo CelularDocument26 pagesCiclo CelularGemma GarciaNo ratings yet
- Esay 3Document6 pagesEsay 3Ismael Delgado SanchezNo ratings yet
- Actividad 7 de Morfo I Adriana BritoDocument11 pagesActividad 7 de Morfo I Adriana BritoAdriana BritoNo ratings yet
- Biologia Tema 11Document5 pagesBiologia Tema 11lucevegonNo ratings yet
- 05-La MitosisDocument10 pages05-La MitosisAnonymous AuBh9cWaNo ratings yet
- Ciclo CelularDocument4 pagesCiclo CelularPINEDA VELASCO DANNA KATHERINENo ratings yet
- Ciclo Celular PDFDocument10 pagesCiclo Celular PDFanita4599No ratings yet
- FecundacionDocument14 pagesFecundacionKatheryne DesiderioNo ratings yet
- Herencia y Variación BiológicaDocument32 pagesHerencia y Variación BiológicaAlbin LopezNo ratings yet
- TRABAJO PRÁCTICO-Reproducción CelularDocument4 pagesTRABAJO PRÁCTICO-Reproducción CelularSilvina RodriguezNo ratings yet
- Mitosis TrabajoDocument14 pagesMitosis TrabajoRogerxVNo ratings yet
- Histología (Tema 2) (Embriologia)Document21 pagesHistología (Tema 2) (Embriologia)Nathalia CasanovaNo ratings yet
- Lectura 5-Division Celular y MeiosisDocument5 pagesLectura 5-Division Celular y Meiosislizzy510No ratings yet
- 2 - 1 2 3 Semana 2013Document20 pages2 - 1 2 3 Semana 2013Micaela QuirogaNo ratings yet
- Inter FaseDocument5 pagesInter Faseandrew1112No ratings yet
- División CelularDocument23 pagesDivisión CelularBeatrizNo ratings yet
- Tercera Parcial Bio9logiaDocument46 pagesTercera Parcial Bio9logiaabelardo llicahua astoNo ratings yet
- Ciclo CelularDocument19 pagesCiclo CelularManuel EduardoNo ratings yet
- Ciclo CelularDocument21 pagesCiclo CelularRushwalker :3No ratings yet
- Meiosis MitosisDocument10 pagesMeiosis Mitosisolga garciaNo ratings yet
- Reporte de Lectura 3Document4 pagesReporte de Lectura 3Luis BrvNo ratings yet
- Tema EmbriologiaDocument18 pagesTema EmbriologiaSofia SirrNo ratings yet
- ProfaseDocument5 pagesProfaseleoNo ratings yet
- FecundacionDocument23 pagesFecundacionMarie ClavijoNo ratings yet
- 05-La Mitosis PDFDocument10 pages05-La Mitosis PDFCapriLaloNo ratings yet
- DIVISIÓN - CELULAR - 5°SEC - 2022 SCDocument20 pagesDIVISIÓN - CELULAR - 5°SEC - 2022 SCJonathanNo ratings yet
- Interfase y Mitosis.Document16 pagesInterfase y Mitosis.Santiago RomeroNo ratings yet
- Asoanato Histo Cat1 Sub Cyd Parte 1Document7 pagesAsoanato Histo Cat1 Sub Cyd Parte 1Virginia DazaNo ratings yet
- ????????????? ???????Document6 pages????????????? ???????paz ávilaNo ratings yet
- Sintesis Reproduccion CelularDocument4 pagesSintesis Reproduccion CelularJohaira GiraldoNo ratings yet
- Material de Estudio BiologíaDocument15 pagesMaterial de Estudio Biologíaadrianoelst15No ratings yet
- Ciclo Celular Puntos de Control Del Ciclo Celular Flujo de Información Genética Ácidos NucleicosDocument29 pagesCiclo Celular Puntos de Control Del Ciclo Celular Flujo de Información Genética Ácidos NucleicosGonzalo LeoNo ratings yet
- Cuestionario Embrio - Docx1Document4 pagesCuestionario Embrio - Docx1Anthony MatosNo ratings yet
- Introducción a la Biología: RESÚMENES UNIVERSITARIOSFrom EverandIntroducción a la Biología: RESÚMENES UNIVERSITARIOSRating: 5 out of 5 stars5/5 (1)
- CarcinogénesisDocument7 pagesCarcinogénesisOs FalNo ratings yet
- 3 Protocolos Histológicos PDFDocument5 pages3 Protocolos Histológicos PDFLamoNo ratings yet
- Alfonso Eduardo - Compendio de EmbriologiaDocument128 pagesAlfonso Eduardo - Compendio de EmbriologiaCarlos_Mata_7428100% (1)
- Diapositivas Del Tejido ÓseoDocument12 pagesDiapositivas Del Tejido ÓseoviruzzNo ratings yet
- Modulo CN 6 B y Qca 2024Document75 pagesModulo CN 6 B y Qca 2024transcopiasNo ratings yet
- Corrimiento Electroforético de DnaDocument8 pagesCorrimiento Electroforético de DnaMiguel Angel Rodas HerreraNo ratings yet
- Evaluacion 8vo Celula Membrana PlasmáticaDocument5 pagesEvaluacion 8vo Celula Membrana Plasmáticajessica gonzalez0% (1)
- Adn y Arn CaracterísticasDocument19 pagesAdn y Arn CaracterísticasDavid Flores100% (1)
- Estreptococos: clasificación, factores de virulencia y enfermedadesDocument182 pagesEstreptococos: clasificación, factores de virulencia y enfermedadesgabriela loarcaNo ratings yet
- 3er Epriodo Ciencias Guia 1teoría-CelularDocument3 pages3er Epriodo Ciencias Guia 1teoría-CelularBetty BuitragoNo ratings yet
- Teoria CelularDocument16 pagesTeoria Celularkalein100% (2)
- ReplicacionDocument28 pagesReplicacionAurelia Gonzales CalleNo ratings yet
- Estructuras y rutas metabólicas bioenergéticasDocument6 pagesEstructuras y rutas metabólicas bioenergéticasOscar ReyesNo ratings yet
- Microbiología 2Document144 pagesMicrobiología 2mix LoveNo ratings yet
- 100 PreguntasDocument15 pages100 Preguntasbeatrixkiddo100No ratings yet
- TincionesDocument44 pagesTincionesLore MgNo ratings yet
- Resumen de Oncogénesis ViralDocument8 pagesResumen de Oncogénesis ViralGeronimo TognottiNo ratings yet
- FG&B Equipo2 Ev1 PDFDocument9 pagesFG&B Equipo2 Ev1 PDFEmilyNo ratings yet
- Histologia Humana Apuntes Histologia PDFDocument54 pagesHistologia Humana Apuntes Histologia PDFRaul Fernando GarzonNo ratings yet
- Estres OxidativoDocument3 pagesEstres OxidativoFatima Quispe CastroNo ratings yet
- La importancia del microscopio en el estudio de las célulasDocument2 pagesLa importancia del microscopio en el estudio de las célulasPatoNo ratings yet
- Informe de Laboratorio 4Document3 pagesInforme de Laboratorio 4SthefiiEnriquez100% (1)
- Producción, Maduración y Diferenciación de LeucocitosDocument41 pagesProducción, Maduración y Diferenciación de LeucocitosTony Tello Enriquez50% (2)
- Resumen 1 ParcialDocument17 pagesResumen 1 ParcialLucía Roque RouviereNo ratings yet
- Epitelio cilios microvellosidadesDocument6 pagesEpitelio cilios microvellosidadesrafael mejiaNo ratings yet
- Cap 036Document4 pagesCap 036Luis Felipe GuzmánNo ratings yet
- Diabetes Melitus-Aula 1Document27 pagesDiabetes Melitus-Aula 1Durão BoavidaNo ratings yet
- DFIFA Sanchez Martin C Capacidad PDFDocument290 pagesDFIFA Sanchez Martin C Capacidad PDFDani LopezNo ratings yet
- Partes de Las Células ProcariontesDocument9 pagesPartes de Las Células Procariontespao3296No ratings yet