You might also like
- Proceedings of the First International Congress of Parasitology: Roma, 21-26 September 1964From EverandProceedings of the First International Congress of Parasitology: Roma, 21-26 September 1964Augusto CorradettiNo ratings yet
- Genetic RecombinationDocument24 pagesGenetic Recombinationkai dollNo ratings yet
- 120 313 Mycology MaielloDocument3 pages120 313 Mycology MaielloBen100% (1)
- Molecular Mechanisms of Insecticide ResistanceDocument11 pagesMolecular Mechanisms of Insecticide ResistancejoharijalinasNo ratings yet
- Chapter - 7: Diversity in Living OrganismsDocument33 pagesChapter - 7: Diversity in Living Organismsuma mishraNo ratings yet
- Parasite Lab 5 Helminths PDF (M)Document18 pagesParasite Lab 5 Helminths PDF (M)Ali RonaldoNo ratings yet
- Systematic Bacteriology: The and The Deeply Branching and PhototrophicDocument18 pagesSystematic Bacteriology: The and The Deeply Branching and PhototrophicMuhammad JamalNo ratings yet
- 1 Ahamd Sarki GumelDocument11 pages1 Ahamd Sarki GumelAhmad Mohammed GumelNo ratings yet
- Modes of Nutrition in Bacteria ?Document7 pagesModes of Nutrition in Bacteria ?Atika ZulfiqarNo ratings yet
- Viruses Ch10.3 7th PDF (Information Obtained From Holt Science and Technology: Life Science. Austin: Holt Rinehart & Winston, 2007. Print.)Document23 pagesViruses Ch10.3 7th PDF (Information Obtained From Holt Science and Technology: Life Science. Austin: Holt Rinehart & Winston, 2007. Print.)thegedusNo ratings yet
- Life Cycle of OedogoniumDocument16 pagesLife Cycle of OedogoniumMuskan Sachdeva 0047No ratings yet
- Bacteria Colony Appearance MorphologyDocument2 pagesBacteria Colony Appearance MorphologyMonique Lopes da SilvaNo ratings yet
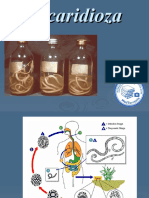
- AscaridiozaDocument25 pagesAscaridiozaLarisa ElenaNo ratings yet
- ChromosomesDocument4 pagesChromosomesعامر جدونNo ratings yet
- CH 11Document18 pagesCH 11dazaikun22032003No ratings yet
- Amebiasis and Giardiasis-FinalDocument25 pagesAmebiasis and Giardiasis-FinalDaniel AtiehNo ratings yet
- Lec 1 Introduction To Host Parasite Interactions MICR2209 2017 PDFDocument31 pagesLec 1 Introduction To Host Parasite Interactions MICR2209 2017 PDFadnankhaNo ratings yet
- BIOB10 Full Study Guide PDFDocument121 pagesBIOB10 Full Study Guide PDFnadaNo ratings yet
- Rickettsia Coxiella: Prof. Khalifa Sifaw GhengheshDocument20 pagesRickettsia Coxiella: Prof. Khalifa Sifaw GhengheshKhalifa Sifaw GhengheshNo ratings yet
- The Proteases PDFDocument86 pagesThe Proteases PDFTotok PurnomoNo ratings yet
- Hyperthermophiles - MicrobiologyDocument15 pagesHyperthermophiles - MicrobiologysathishNo ratings yet
- LysosomesDocument9 pagesLysosomesapi-318784000No ratings yet
- RNA Editing in TrypanosomesDocument31 pagesRNA Editing in TrypanosomesAarthi VijayNo ratings yet
- Microbial Genetics & MutationDocument49 pagesMicrobial Genetics & MutationRenz Francis SasaNo ratings yet
- Submitted by Aiswarya V 1St MSC Zoology Roll Number 3301Document32 pagesSubmitted by Aiswarya V 1St MSC Zoology Roll Number 3301PES PEOPLENo ratings yet
- Cell Profile Report - Nisal Karawita - Nlkar1Document12 pagesCell Profile Report - Nisal Karawita - Nlkar1Lochana Karawita100% (1)
- Parasites & People - Host Parasite Relationship - RumalaDocument40 pagesParasites & People - Host Parasite Relationship - RumalamicroperadeniyaNo ratings yet
- Genome Size and Structure, BacterialDocument7 pagesGenome Size and Structure, BacterialCINDY NATHALIA GARCIA AGUDELONo ratings yet
- Fate Maps: Course: B.Sc. (H) Zoology VI Semester Paper: Developmental Biology Faculty: Dr. Priya GoelDocument17 pagesFate Maps: Course: B.Sc. (H) Zoology VI Semester Paper: Developmental Biology Faculty: Dr. Priya GoelSusmita PalNo ratings yet
- Molecular Identification of Bacteria IIII 2015Document121 pagesMolecular Identification of Bacteria IIII 2015Mohammed MohsenNo ratings yet
- Histone ModificationsDocument61 pagesHistone ModificationsNitin BagraNo ratings yet
- Blast Vs FastaDocument3 pagesBlast Vs FastaAhmad Faraz KhanNo ratings yet
- Le Stress OxydantDocument8 pagesLe Stress Oxydantbiotech_khaledNo ratings yet
- Ferns & Fern AlliesDocument7 pagesFerns & Fern AlliesNicaRejusoNo ratings yet
- Theory and Strategy in HistochemistryDocument581 pagesTheory and Strategy in HistochemistryNoa GoyanesNo ratings yet
- Anatomy of Flowering PlantsDocument33 pagesAnatomy of Flowering PlantsRaichal P Biju100% (1)
- Ultracentrifugation: June 2010Document8 pagesUltracentrifugation: June 2010Nayak Alok RanjanNo ratings yet
- BRYOPHYTES AND EARLY VASCULAR PLANTSDocument101 pagesBRYOPHYTES AND EARLY VASCULAR PLANTSRyan Merza100% (1)
- 1# Lectr - Botany - 1 Concepts of Botany An Introducton To Plant BiologyDocument33 pages1# Lectr - Botany - 1 Concepts of Botany An Introducton To Plant BiologyEsra Yuliana manalu100% (1)
- Lysosomes: Cell BiologyDocument20 pagesLysosomes: Cell BiologyBio SciencesNo ratings yet
- Fungi Morphology and Classification ExplainedDocument37 pagesFungi Morphology and Classification Explainedjihan faradhilaNo ratings yet
- Notes - Growth and DevelopmentDocument4 pagesNotes - Growth and DevelopmentNathan mwapeNo ratings yet
- Structure of Prion: The Normal Prion Structure Is Characterized by Four Helices, Denoted As PRPCDocument14 pagesStructure of Prion: The Normal Prion Structure Is Characterized by Four Helices, Denoted As PRPCMatthew WilsonNo ratings yet
- 1 - Introduction To ParasitologyDocument60 pages1 - Introduction To ParasitologyThesa TagalogNo ratings yet
- Clonorchis SinensisDocument20 pagesClonorchis SinensisDedy SavradinataNo ratings yet
- Class MusciDocument18 pagesClass MusciElaine BriososNo ratings yet
- ColeochaeteDocument24 pagesColeochaeteC.V. Kalpana100% (1)
- Bacteria Nutrition RequirementsDocument14 pagesBacteria Nutrition RequirementsFuad Hasan Pranto 1921147049No ratings yet
- Phytosequestration Mechanisms in RhizofiltrationDocument42 pagesPhytosequestration Mechanisms in RhizofiltrationSeroKeretaMasaroWidiarNo ratings yet
- MM North BlotDocument30 pagesMM North Blotapi-320465752No ratings yet
- Senior I-Search Paper - Naomi Williams Parts 1-4Document17 pagesSenior I-Search Paper - Naomi Williams Parts 1-4api-526305574No ratings yet
- Botanica Sistematica 2Document44 pagesBotanica Sistematica 2Maya PelinNo ratings yet
- Development of AmphioxusDocument59 pagesDevelopment of AmphioxusDyah Ayu PuspitasariNo ratings yet
- Toxoplasma gondii: An Obligate ParasiteDocument7 pagesToxoplasma gondii: An Obligate ParasiteDedy SavradinataNo ratings yet
- Relationship Between Dna, Gene ChromosomeDocument31 pagesRelationship Between Dna, Gene ChromosomemaimonsabiNo ratings yet
- Frog Embryo DevelopmentDocument36 pagesFrog Embryo DevelopmentMomina ButtNo ratings yet
- Insect Physiology: Digestive SystemDocument8 pagesInsect Physiology: Digestive SystemShyamol Bose100% (1)
- Solution Sheets AcetoorceinDocument1 pageSolution Sheets AcetoorceinsaputraalamNo ratings yet
- Reproduction in AlgaeDocument57 pagesReproduction in AlgaeNarendra Kumar100% (1)
- ArchaeaDocument9 pagesArchaeaUjjawal KumarNo ratings yet
- Antibiotic Production by RhizobacteriaDocument29 pagesAntibiotic Production by RhizobacteriaDr. Farokh Rokhbakhsh-ZaminNo ratings yet
- Cytotoxic Activity of Medicinal Plants Used in Iranian Traditional Medicine On Two Strains of Saccharomyces CerevisiaeDocument3 pagesCytotoxic Activity of Medicinal Plants Used in Iranian Traditional Medicine On Two Strains of Saccharomyces CerevisiaeDr. Farokh Rokhbakhsh-ZaminNo ratings yet
- Characterization of PGP Traits of Acinetobacter Species Isolated From Rhizosphere of Pennisetum GlaucumDocument1 pageCharacterization of PGP Traits of Acinetobacter Species Isolated From Rhizosphere of Pennisetum GlaucumDr. Farokh Rokhbakhsh-ZaminNo ratings yet
- Biofilm Formation by Acinetobacter Baumannii Strains Isolated From UTI & Urinary CathetersDocument2 pagesBiofilm Formation by Acinetobacter Baumannii Strains Isolated From UTI & Urinary CathetersDr. Farokh Rokhbakhsh-ZaminNo ratings yet
- Isolation of Acinetobacter Spp. From Rhizosphere of Crop PlantsDocument1 pageIsolation of Acinetobacter Spp. From Rhizosphere of Crop PlantsDr. Farokh Rokhbakhsh-ZaminNo ratings yet
- Agricultural Biotechnology - BiofertilizersDocument38 pagesAgricultural Biotechnology - BiofertilizersDr. Farokh Rokhbakhsh-Zamin100% (17)
- 6th Central Pay Commission Salary CalculatorDocument15 pages6th Central Pay Commission Salary Calculatorrakhonde100% (436)
- A Study On Consumer Behavior in Auro Food (True Biscuits) PVT LTDDocument52 pagesA Study On Consumer Behavior in Auro Food (True Biscuits) PVT LTDRoyal Projects100% (1)
- The Islam Question - Should I Become A Muslim?Document189 pagesThe Islam Question - Should I Become A Muslim?Aorounga100% (1)
- Modelling of Induction Motor PDFDocument42 pagesModelling of Induction Motor PDFsureshNo ratings yet
- Vinzenz Hediger, Patrick Vonderau - Films That Work - Industrial Film and The Productivity of Media (Film Culture in Transition) (2009)Document496 pagesVinzenz Hediger, Patrick Vonderau - Films That Work - Industrial Film and The Productivity of Media (Film Culture in Transition) (2009)Arlindo Rebechi JuniorNo ratings yet
- Sexual Self PDFDocument23 pagesSexual Self PDFEden Faith Aggalao100% (1)
- Twin-Field Quantum Key Distribution Without Optical Frequency DisseminationDocument8 pagesTwin-Field Quantum Key Distribution Without Optical Frequency DisseminationHareesh PanakkalNo ratings yet
- Web Search - One People's Public Trust 1776 UCCDocument28 pagesWeb Search - One People's Public Trust 1776 UCCVincent J. CataldiNo ratings yet
- Bible Study RisksDocument6 pagesBible Study RisksVincentNo ratings yet
- Tle-Bpp 8-Q1-M18Document14 pagesTle-Bpp 8-Q1-M18Michelle LlanesNo ratings yet
- Code of Conduct GuidanceDocument17 pagesCode of Conduct GuidanceMuhammad RidwanNo ratings yet
- Reconsidering Puerto Rico's Status After 116 Years of Colonial RuleDocument3 pagesReconsidering Puerto Rico's Status After 116 Years of Colonial RuleHéctor Iván Arroyo-SierraNo ratings yet
- Chapter 1. Introduction To TCPIP NetworkingDocument15 pagesChapter 1. Introduction To TCPIP NetworkingPoojitha NagarajaNo ratings yet
- What Music Really Means To ChildrenDocument5 pagesWhat Music Really Means To ChildrenMara Sofia ValenteNo ratings yet
- Why Research Is Important in The BusinessDocument2 pagesWhy Research Is Important in The BusinessBricx BalerosNo ratings yet
- Lung BiopsyDocument8 pagesLung BiopsySiya PatilNo ratings yet
- Personal Weaknesses ListDocument3 pagesPersonal Weaknesses ListKinga SzászNo ratings yet
- Intrinsic Resistance and Unusual Phenotypes Tables v3.2 20200225Document12 pagesIntrinsic Resistance and Unusual Phenotypes Tables v3.2 20200225Roy MontoyaNo ratings yet
- PIC16 F 1619Document594 pagesPIC16 F 1619Francisco Martinez AlemanNo ratings yet
- Week 1 Amanda CeresaDocument2 pagesWeek 1 Amanda CeresaAmanda CeresaNo ratings yet
- Scientech 2502ADocument2 pagesScientech 2502Aashutosh kumarNo ratings yet
- Jolly Phonics Teaching Reading and WritingDocument6 pagesJolly Phonics Teaching Reading and Writingmarcela33j5086100% (1)
- Engineering Economy Course SyllabusDocument11 pagesEngineering Economy Course Syllabuschatter boxNo ratings yet
- Curriculum VitaeDocument8 pagesCurriculum VitaeChristine LuarcaNo ratings yet
- CBCP Monitor Vol. 17 No. 9Document20 pagesCBCP Monitor Vol. 17 No. 9Areopagus Communications, Inc.No ratings yet
- Intermediate Reading Comprehension Test 03Document5 pagesIntermediate Reading Comprehension Test 03MZNo ratings yet
- Bombardier CityfloDocument14 pagesBombardier CityfloBiju KmNo ratings yet
- Semi-Detailed Lesson Plan Template: Pagsanjan Intergrated National High School 8Document3 pagesSemi-Detailed Lesson Plan Template: Pagsanjan Intergrated National High School 8Mae Ganate RoblesNo ratings yet
- Module 2 - Content and Contextual Analysis of Selected Primary andDocument41 pagesModule 2 - Content and Contextual Analysis of Selected Primary andAngelica CaldeoNo ratings yet
- OEO105020 LTE ERAN2.2 Connection Management Feature ISSUE 1.00Document52 pagesOEO105020 LTE ERAN2.2 Connection Management Feature ISSUE 1.00Daniel YulistianNo ratings yet
- Understanding Deuteronomy On Its Own TermsDocument5 pagesUnderstanding Deuteronomy On Its Own TermsAlberto RodriguesNo ratings yet