You might also like
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- 7th Grade ScienceDocument149 pages7th Grade ScienceSirleny100% (4)
- 7th Grade ScienceDocument149 pages7th Grade ScienceSirleny100% (4)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Earth Science PDFDocument394 pagesEarth Science PDFnfalkdrf alkfalk100% (2)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (890)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Grade5 Science Interactive Text PDFDocument208 pagesGrade5 Science Interactive Text PDFrheeya08100% (2)
- Cambridge Checkpoint Science Coursebook 8pdf PDFDocument190 pagesCambridge Checkpoint Science Coursebook 8pdf PDFHùng Võ Hoàng86% (58)
- Essential Molecular Biology - A Practical Approach Volume I (Practical Approach Series) (2nd Edition) PDFDocument261 pagesEssential Molecular Biology - A Practical Approach Volume I (Practical Approach Series) (2nd Edition) PDFAvinash100% (1)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- The MycotaDocument407 pagesThe MycotaHa Bui100% (1)
- California Earth Science Grade6 ReadingWritingDocument176 pagesCalifornia Earth Science Grade6 ReadingWritingviswamanojNo ratings yet
- Gr6 Science Activity PDFDocument278 pagesGr6 Science Activity PDFHa BuiNo ratings yet
- sc08 A6icr Web PDFDocument232 pagessc08 A6icr Web PDFMensah Gbeassor100% (1)
- Molecular Biology and Diagnostics Course DeliveryDocument34 pagesMolecular Biology and Diagnostics Course DeliveryAbigail Laus100% (2)
- Gr5 Science Test Prep PDFDocument73 pagesGr5 Science Test Prep PDFHa BuiNo ratings yet
- CH 05Document24 pagesCH 05filippo100% (3)
- The Human Genome ProjectDocument22 pagesThe Human Genome ProjectAashianaThiyam100% (1)
- Mol 214 Lab Report #1Document5 pagesMol 214 Lab Report #1Samuel RobertsNo ratings yet
- Bioethanol - The Fuel of Tomorrow From The Residue of TodayDocument8 pagesBioethanol - The Fuel of Tomorrow From The Residue of TodaytvdaihaiNo ratings yet
- Wastewater Treatment A Model System For Microbial EcologyDocument7 pagesWastewater Treatment A Model System For Microbial EcologyHa BuiNo ratings yet
- Apoptosis and Cell ProliferationDocument143 pagesApoptosis and Cell ProliferationShruthi SunderrajanNo ratings yet
- Bacterial Sensing and Signaling-Karger (2009)Document239 pagesBacterial Sensing and Signaling-Karger (2009)Ha BuiNo ratings yet
- Bioethanol - The Fuel of Tomorrow From The Residue of TodayDocument8 pagesBioethanol - The Fuel of Tomorrow From The Residue of TodaytvdaihaiNo ratings yet
- Biology A2 Snab CourseworkDocument120 pagesBiology A2 Snab CourseworkIshwarya SivakumarNo ratings yet
- RAPD Markers For Identifying Oil Palm Jacq.) Parental Varieties & and The HybridDocument7 pagesRAPD Markers For Identifying Oil Palm Jacq.) Parental Varieties & and The HybridRamNo ratings yet
- Analytical Biochemistry 1990 Engelke Taq PurificationDocument5 pagesAnalytical Biochemistry 1990 Engelke Taq PurificationPablo L. Cossio RodriguezNo ratings yet
- Evaluation of The Anti Genotoxicity and Growth Performance Impacts of Green Algae On M CephalusDocument12 pagesEvaluation of The Anti Genotoxicity and Growth Performance Impacts of Green Algae On M CephalusJuan FranciaNo ratings yet
- Types of BlottingDocument7 pagesTypes of Blottingporkodi sudhaNo ratings yet
- Standard Operating Procedures MycologyDocument48 pagesStandard Operating Procedures MycologyvishnupgiNo ratings yet
- LS23L S12 Syllabus Week by Week GridDocument8 pagesLS23L S12 Syllabus Week by Week Gridkrngirl10No ratings yet
- CAIE Chemistry A-Level: 34: Nitrogen CompoundsDocument9 pagesCAIE Chemistry A-Level: 34: Nitrogen CompoundsahumanbeinginearthNo ratings yet
- Amino AcidDocument24 pagesAmino AcidAdisa YellowNo ratings yet
- Lab 2 ds180 Genotyping LabDocument7 pagesLab 2 ds180 Genotyping Labapi-342081300No ratings yet
- Human Genome ProjectDocument42 pagesHuman Genome ProjectGopikagopinathan RajeshNo ratings yet
- Ez-Pcr Mycoplasma Test Kit: Instructions For UseDocument2 pagesEz-Pcr Mycoplasma Test Kit: Instructions For Usenauval mangantjoNo ratings yet
- Recognized That Each Individual Has A Unique Pattern of MinisatellitesDocument4 pagesRecognized That Each Individual Has A Unique Pattern of Minisatelliteskeijanne marie mendozaNo ratings yet
- Web SitesDocument41 pagesWeb Sitesเกมกวี MedicalStudentNo ratings yet
- Gel electrophoresis separates proteins by sizeDocument4 pagesGel electrophoresis separates proteins by sizeRommel BauzaNo ratings yet
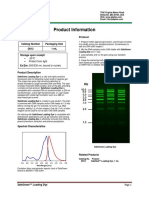
- Product Information: Safegreen ™ Loading Dye Catalog Number Packaging Size D012 1 ML ProtocolDocument1 pageProduct Information: Safegreen ™ Loading Dye Catalog Number Packaging Size D012 1 ML ProtocolkokoNo ratings yet
- Plasmid LabDocument10 pagesPlasmid LabAhmed J AlhindaweNo ratings yet
- Agarose Gel ElectrophoresisDocument10 pagesAgarose Gel Electrophoresisvenkat satyaNo ratings yet
- Bio Module 1Document43 pagesBio Module 1XdxdNo ratings yet
- Recombinant DNA cloning techniquesDocument101 pagesRecombinant DNA cloning techniquesdolt2111No ratings yet
- Dirofilaria Immitis in Bulgaria The First Genetic Baseline Data and An Overview of The Current StatusDocument8 pagesDirofilaria Immitis in Bulgaria The First Genetic Baseline Data and An Overview of The Current StatusHelenaNitoNo ratings yet
- Gel ElectroiphoresisDocument22 pagesGel ElectroiphoresisAqsa MazharNo ratings yet
- Basic Techniques To Manipulate Genetic MaterialDocument7 pagesBasic Techniques To Manipulate Genetic MaterialRogelio Arellano LawayanNo ratings yet
- General Catalog PT. Genecraft LabsDocument33 pagesGeneral Catalog PT. Genecraft LabsbukapintuNo ratings yet
- ELECTROPHORESISDocument19 pagesELECTROPHORESISAmit SahNo ratings yet