You might also like
- Fluorescence Microscopy: Super-Resolution and other Novel TechniquesFrom EverandFluorescence Microscopy: Super-Resolution and other Novel TechniquesAnda CorneaNo ratings yet
- Electrophysiology Measurements for Studying Neural InterfacesFrom EverandElectrophysiology Measurements for Studying Neural InterfacesNo ratings yet
- Oe 13 23 9361Document13 pagesOe 13 23 9361Aggelos StefopoulosNo ratings yet
- 1 s2.0 S0022073606001282 MainDocument5 pages1 s2.0 S0022073606001282 Main6pb4rwpk45No ratings yet
- Ert 445Document8 pagesErt 445Mirai AkihitoNo ratings yet
- PNAS 2001 Basché 10527 8Document2 pagesPNAS 2001 Basché 10527 8Jazmín Gonzales TovarNo ratings yet
- Stimulated Emission Depletion (STED) Nanoscopy of A Fluorescent Protein-Labeled Organelle Inside A Living CellDocument6 pagesStimulated Emission Depletion (STED) Nanoscopy of A Fluorescent Protein-Labeled Organelle Inside A Living CellAMNo ratings yet
- Meng 2022 J. Opt. 24 054004Document8 pagesMeng 2022 J. Opt. 24 054004Ozan KarNo ratings yet
- 01 EsselingDocument6 pages01 EsselingRakesh KhannaNo ratings yet
- Optical OptogeneticsDocument22 pagesOptical OptogeneticshisenknNo ratings yet
- Correlative microscopy 02Document13 pagesCorrelative microscopy 02UNIG Aluana SantanaNo ratings yet
- Surface Plasmon Resonance ThesisDocument6 pagesSurface Plasmon Resonance Thesisbsna8p6k100% (2)
- Burel Etal Development 145 Dev160879 2018 RedDocument22 pagesBurel Etal Development 145 Dev160879 2018 RedCarlla Araújo Guarani KaiowáNo ratings yet
- Biophotons PDFDocument22 pagesBiophotons PDFrafaelNo ratings yet
- Biomedical Optical Phase Microscopy and NanoscopyFrom EverandBiomedical Optical Phase Microscopy and NanoscopyNatan T. ShakedNo ratings yet
- Nanohole Biosensor-Origin and Application As Multiplex Biosensing PlatformDocument3 pagesNanohole Biosensor-Origin and Application As Multiplex Biosensing PlatformShibsekhar RoyNo ratings yet
- Emission of Mitochondrial BiophotonsDocument24 pagesEmission of Mitochondrial BiophotonsGabrielNo ratings yet
- Corelative Fluorescence and Electron MicrosDocument10 pagesCorelative Fluorescence and Electron MicrosEmma StepinacNo ratings yet
- Brain Atrophy of An Alzheimer Disease Affected PatientDocument22 pagesBrain Atrophy of An Alzheimer Disease Affected PatientEnamulduNo ratings yet
- Surface Plasmon Resonance DissertationDocument6 pagesSurface Plasmon Resonance DissertationCollegePaperWritingServiceReviewsSingapore100% (1)
- A Dom 201400472Document7 pagesA Dom 201400472aishuvc1822No ratings yet
- Transmission by Solitary Waves in Biological Systems: LeninskyDocument11 pagesTransmission by Solitary Waves in Biological Systems: LeninskyLejirbag CevardazNo ratings yet
- TMP 51 BDocument7 pagesTMP 51 BFrontiersNo ratings yet
- New Crystal in The Pineal Gland: Characterization and Potential Role in Electromechano-TransductionDocument4 pagesNew Crystal in The Pineal Gland: Characterization and Potential Role in Electromechano-TransductionMarkoff ChaneyNo ratings yet
- Biointerface Characterization by Advanced IR SpectroscopyFrom EverandBiointerface Characterization by Advanced IR SpectroscopyC.-M. PradierRating: 5 out of 5 stars5/5 (1)
- STEM3Document6 pagesSTEM3UNIG Aluana SantanaNo ratings yet
- The Study of Electrokinetic Properties of Buccal EpitheliumDocument5 pagesThe Study of Electrokinetic Properties of Buccal Epitheliumgeorgbell5No ratings yet
- Nanoscale Laminin Coating Modulates Cortical Scarring Response Around Implanted Silicon Microelectrode ArraysDocument11 pagesNanoscale Laminin Coating Modulates Cortical Scarring Response Around Implanted Silicon Microelectrode ArraysHutsDMNo ratings yet
- Conceitos de Microscopia de Super Resolução em FluorescênciaDocument13 pagesConceitos de Microscopia de Super Resolução em FluorescênciaUNIG Aluana SantanaNo ratings yet
- A Simple Method For Quantitating Confocal Fluorescent ImagesDocument21 pagesA Simple Method For Quantitating Confocal Fluorescent Imagesyylf2016No ratings yet
- SRRFDocument6 pagesSRRFhdgkorea19No ratings yet
- Transmission Electron Microscopy Research PapersDocument4 pagesTransmission Electron Microscopy Research PapersgvzexnzhNo ratings yet
- Group 5 Human AnaPhy With Pathophysiology QuestionsDocument2 pagesGroup 5 Human AnaPhy With Pathophysiology QuestionsJuneyale PadillaNo ratings yet
- 2019 FREIA ReportDocument18 pages2019 FREIA ReportAlenNo ratings yet
- MOF-photodetector-AdvSci - 2021Document8 pagesMOF-photodetector-AdvSci - 2021du yixuanNo ratings yet
- Presented by Guided By: Ms - Arpana Pancholi M.SC Biotechnology I Semester Mr. Obulesu M Lect. Deparment of BiotechDocument30 pagesPresented by Guided By: Ms - Arpana Pancholi M.SC Biotechnology I Semester Mr. Obulesu M Lect. Deparment of Biotecharpana4317No ratings yet
- Combined scanning probe and total internal reflection fluorescence microscopyDocument9 pagesCombined scanning probe and total internal reflection fluorescence microscopyNguyễn ThuNo ratings yet
- 2012 - Lucas, Riedo - Invited Review Article Combining Scanning Probe Microscopy With Optical Spectroscopy For Applications in Biology ADocument37 pages2012 - Lucas, Riedo - Invited Review Article Combining Scanning Probe Microscopy With Optical Spectroscopy For Applications in Biology AClaudio BiaginiNo ratings yet
- HighOrderAberrations02-Zivanov 2020Document15 pagesHighOrderAberrations02-Zivanov 2020Tongyi DouNo ratings yet
- Europhotonics Emjd Doctorate Program PHD Projects 2011 - 2014Document7 pagesEurophotonics Emjd Doctorate Program PHD Projects 2011 - 2014Preetha AlbertNo ratings yet
- Imaging The Spatial Evolution of Degradation in Perovskite/Si Tandem Solar Cells After Exposure To Humid AirDocument6 pagesImaging The Spatial Evolution of Degradation in Perovskite/Si Tandem Solar Cells After Exposure To Humid AirDiego FrizterNo ratings yet
- Conceitos de Microscopia de FluorescênciaDocument25 pagesConceitos de Microscopia de FluorescênciaUNIG Aluana SantanaNo ratings yet
- Bio-Inspired Polarization Imaging Sensors: From Circuits and Optics To Signal Processing Algorithms and Biomedical ApplicationsDocument16 pagesBio-Inspired Polarization Imaging Sensors: From Circuits and Optics To Signal Processing Algorithms and Biomedical ApplicationsJaya SwaroopNo ratings yet
- Mechanisms of Femtosecond Laser Nanosurgery of Cells and Tissues - VogelDocument33 pagesMechanisms of Femtosecond Laser Nanosurgery of Cells and Tissues - VogelBiswajit MishraNo ratings yet
- Lechasseur2011 PDFDocument9 pagesLechasseur2011 PDFMd. HasanuzzamanNo ratings yet
- Techniques in Cell and Molecular BiologyDocument40 pagesTechniques in Cell and Molecular BiologyWai Kwong ChiuNo ratings yet
- Mapping Neuronal DiversityDocument2 pagesMapping Neuronal DiversityJohn MistryNo ratings yet
- On-Chip Capacitance Sensing For Cell Monitoring ApplicationsDocument8 pagesOn-Chip Capacitance Sensing For Cell Monitoring ApplicationsAlex WongNo ratings yet
- Development of Hardware-Software Microscopy Complex For The Study of Buccal Epithelial CellsDocument6 pagesDevelopment of Hardware-Software Microscopy Complex For The Study of Buccal Epithelial Cellsgeorgbell5No ratings yet
- Journal Name: CommunicationDocument3 pagesJournal Name: CommunicationzuopengxiangNo ratings yet
- Recent Progress in Optical Sensors For Biomedical DiagnosticsDocument4 pagesRecent Progress in Optical Sensors For Biomedical DiagnosticsFyaNo ratings yet
- ElectrophysiologyDocument32 pagesElectrophysiologyAbdelrehim SirajNo ratings yet
- ElectrophysiologyDocument32 pagesElectrophysiologyAbdelrehim Siraj100% (7)
- S41467-018-04116-9-Articol TezaDocument10 pagesS41467-018-04116-9-Articol TezaElena AnghelNo ratings yet
- OCT Imaging in Glaucoma: A guide for practitionersFrom EverandOCT Imaging in Glaucoma: A guide for practitionersKi Ho ParkNo ratings yet
- Term Paper On Transmission Electron MicroscopeDocument7 pagesTerm Paper On Transmission Electron Microscopeafdtveepo100% (1)
- TMP 610 CDocument6 pagesTMP 610 CFrontiersNo ratings yet
- Beyond Fluorescence: News & ViewsDocument2 pagesBeyond Fluorescence: News & Viewstramvu93No ratings yet
- JurnalDocument6 pagesJurnalNur AnindyaNo ratings yet
- Nihms 789951 PDFDocument21 pagesNihms 789951 PDFNur AnindyaNo ratings yet
- OHSSDocument4 pagesOHSSNur AnindyaNo ratings yet
- Infertility and Women: Hormones & YouDocument1 pageInfertility and Women: Hormones & YouNur AnindyaNo ratings yet
- Prevention and Treatment of Moderate and Severe Ovarian Hyperstimulation Syndrome NoprintDocument14 pagesPrevention and Treatment of Moderate and Severe Ovarian Hyperstimulation Syndrome NoprintDo le QuangNo ratings yet
- JurnalDocument13 pagesJurnalNur AnindyaNo ratings yet
- Purine and Pyrimidine Contents of Desoxypentose Nucleic AcidsDocument14 pagesPurine and Pyrimidine Contents of Desoxypentose Nucleic AcidsNur AnindyaNo ratings yet
- Ovarian Hyperstimulation Syndrome Diagnosis and ManagementDocument21 pagesOvarian Hyperstimulation Syndrome Diagnosis and ManagementNur AnindyaNo ratings yet
- JurnalDocument13 pagesJurnalNur AnindyaNo ratings yet
- Female Reproductive Tract and Mammary Disorders Caused by Endocrine DisruptorsDocument4 pagesFemale Reproductive Tract and Mammary Disorders Caused by Endocrine DisruptorsNur AnindyaNo ratings yet
- 4Document19 pages4Nur AnindyaNo ratings yet
- Gene Expression and Maturation Evaluation of Sheep Oocytes Cultured in Medium Supplemented With Natural Antioxidant SourceDocument13 pagesGene Expression and Maturation Evaluation of Sheep Oocytes Cultured in Medium Supplemented With Natural Antioxidant SourceErik Angga SaputraNo ratings yet
- Previews: DNA Made of Purines OnlyDocument3 pagesPreviews: DNA Made of Purines OnlyNur AnindyaNo ratings yet
- JurnalDocument13 pagesJurnalNur AnindyaNo ratings yet
- Wespes Et Al. Eur Urol 2006-49-806 815. EAU Guidelines On Erectile Dysfunction An UpdateDocument10 pagesWespes Et Al. Eur Urol 2006-49-806 815. EAU Guidelines On Erectile Dysfunction An UpdateNur AnindyaNo ratings yet
- 14 Male Sexual Dysfunction LR1Document38 pages14 Male Sexual Dysfunction LR1Nur AnindyaNo ratings yet
- Slurry PropertiesDocument5 pagesSlurry PropertiesJuan Pablo ApazaNo ratings yet
- P7 Revision QuestionsDocument4 pagesP7 Revision Questionsapi-27344426No ratings yet
- Millman HakyaisDocument342 pagesMillman HakyaisBharat BadhanNo ratings yet
- NPTEL Phase II - Mechanical Engineering - Engineering Fracture MechanicsDocument5 pagesNPTEL Phase II - Mechanical Engineering - Engineering Fracture MechanicsSoupramanien KathirvelouNo ratings yet
- Linear Integrated Circuit: Low Frequency Power AmplifierDocument7 pagesLinear Integrated Circuit: Low Frequency Power AmplifierRenzo Fernandez HurtadoNo ratings yet
- Quantum Field TheoryDocument3 pagesQuantum Field TheoryvajrahastaNo ratings yet
- A Review of Residual Stress Measurement MethodsDocument45 pagesA Review of Residual Stress Measurement MethodsCarlos MartinsNo ratings yet
- Department of Applied Physics Question Bank-I Semester PH6151-Engineering Physics - I UNIT-II - Properties of Matter and Thermal PhysicsDocument4 pagesDepartment of Applied Physics Question Bank-I Semester PH6151-Engineering Physics - I UNIT-II - Properties of Matter and Thermal PhysicsJohn Ernie SelmarNo ratings yet
- Daniel Neuhauser and Roi Baer - A Two-Grid Time-Dependent Formalism For The Maxwell EquationDocument10 pagesDaniel Neuhauser and Roi Baer - A Two-Grid Time-Dependent Formalism For The Maxwell EquationPrem_SwiftNo ratings yet
- Physics - Study Guide - Tim Kirk - Oxford 2014 (Dragged) 5Document13 pagesPhysics - Study Guide - Tim Kirk - Oxford 2014 (Dragged) 5Trúc HồNo ratings yet
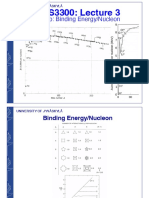
- Binding Energy/Nucleon and Semi-Empirical Mass FormulaDocument28 pagesBinding Energy/Nucleon and Semi-Empirical Mass FormulaKeijo SalonenNo ratings yet
- Advanced Solid State Physics Lab ReportDocument32 pagesAdvanced Solid State Physics Lab ReportLakshay BhardwajNo ratings yet
- Uplift With Moment CalculationDocument4 pagesUplift With Moment CalculationdennykvgNo ratings yet
- Guia Gases 1 1Document11 pagesGuia Gases 1 1FeriaNo ratings yet
- Assignment 2Document4 pagesAssignment 2blackbeauty14No ratings yet
- Lab Reports Mechanics, Heat & VibrationsDocument79 pagesLab Reports Mechanics, Heat & VibrationsNabeel AhmadNo ratings yet
- Concept Strengthening Sheet (CSS-05) Based On AIATS-05 (CF+OYM) - ChemistryDocument4 pagesConcept Strengthening Sheet (CSS-05) Based On AIATS-05 (CF+OYM) - Chemistryshakuntla6413No ratings yet
- Wheatston BridgeDocument12 pagesWheatston BridgeZaid Al-AliNo ratings yet
- Chapter 19Document30 pagesChapter 19d4rkne5sNo ratings yet
- UntitledDocument383 pagesUntitledOppo Neo7No ratings yet
- Mazas de Ruedas Larrique RulemanesDocument23 pagesMazas de Ruedas Larrique RulemanesPaisaje SilenciosoNo ratings yet
- Electromagnetic FieldDocument6 pagesElectromagnetic Fieldalokesh1982No ratings yet
- Transformer Testing Purchase OrderDocument2 pagesTransformer Testing Purchase Orderabdulyunus_amirNo ratings yet
- Equipment Lab4 BBL - BoilerType2Document917 pagesEquipment Lab4 BBL - BoilerType2umairsaleem606No ratings yet
- Magnetic Forces, Materials, and Devices: Islamic University of Gaza - Palestine 2012Document9 pagesMagnetic Forces, Materials, and Devices: Islamic University of Gaza - Palestine 2012Amupolo CarlyNo ratings yet
- Jurong Junior College: Physics 9646/2Document22 pagesJurong Junior College: Physics 9646/2Chen ZhihaoNo ratings yet
- H2 Math Paper 1 SolutionsDocument30 pagesH2 Math Paper 1 SolutionsHANSNo ratings yet
- Hydrostatic ForcesDocument27 pagesHydrostatic ForcesSumit SahrawatNo ratings yet
- TorsionDocument12 pagesTorsionredz00No ratings yet
- Experiment 09: Preparation and Characterization of Alkene Tubog, Jalil Venice B., Marilyn MirandaDocument2 pagesExperiment 09: Preparation and Characterization of Alkene Tubog, Jalil Venice B., Marilyn MirandaJalil Venice Bernal TubogNo ratings yet