You might also like
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (890)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- M 371Document14 pagesM 371Anonymous h0DxuJTNo ratings yet
- Radiology ManualDocument24 pagesRadiology ManualQuality Manager100% (2)
- Cancer vs CarcinogensDocument20 pagesCancer vs CarcinogensChelzy Catabas100% (2)
- HT Age Based PDFDocument8 pagesHT Age Based PDFDoni TrinandaNo ratings yet
- Comparison Between Traditional 2-Dimensional Cephalometry and A 3-Dimensional Approach On Human Dry SkullsDocument13 pagesComparison Between Traditional 2-Dimensional Cephalometry and A 3-Dimensional Approach On Human Dry SkullsMarllury PabonNo ratings yet
- CVA Case PresentationDocument76 pagesCVA Case PresentationSi VeekeeNo ratings yet
- ONCOLOGY Board Exam QuestionDocument11 pagesONCOLOGY Board Exam QuestionEJ S Molina93% (15)
- Manual de Operacion JuniperDocument194 pagesManual de Operacion JuniperPepo100% (1)
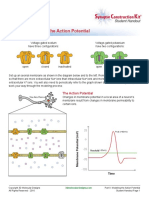
- Part II: Modeling The Action Potential: Student HandoutDocument4 pagesPart II: Modeling The Action Potential: Student HandoutEthan GrangerNo ratings yet
- Answer To VMD Medical Imaging CenterDocument2 pagesAnswer To VMD Medical Imaging CenterPragathi SundarNo ratings yet
- TG40Document39 pagesTG40jiar001No ratings yet
- 2010 Panagiotaki Et Al. Lecture Notes in Computer Science PDFDocument8 pages2010 Panagiotaki Et Al. Lecture Notes in Computer Science PDFAjayaKumarKavalaNo ratings yet
- Basic Principles of MagneticDocument8 pagesBasic Principles of MagneticIon CîmpeanuNo ratings yet
- Radiology Clinics of North America Cardiac ImagingDocument200 pagesRadiology Clinics of North America Cardiac ImagingArie Dwisetyo Gusti Laksono100% (2)
- Neonatal Neurosonography: A Pictorial Essay: EuroradiologyDocument13 pagesNeonatal Neurosonography: A Pictorial Essay: EuroradiologyAnom RahartoNo ratings yet
- Answer All MCQ Questions From No. 1 To 50. Each Question Will Carry 2 MarksDocument4 pagesAnswer All MCQ Questions From No. 1 To 50. Each Question Will Carry 2 MarksRick DasNo ratings yet
- Ao ADocument6 pagesAo AarbazNo ratings yet
- Lecture 1Document21 pagesLecture 1Philip_830No ratings yet
- Physics of Medical X-Ray Imaging (1) Chapter 10: Figure 10.1. Digital Radiographic System DesignDocument21 pagesPhysics of Medical X-Ray Imaging (1) Chapter 10: Figure 10.1. Digital Radiographic System DesignNguyen Cong HuynhNo ratings yet
- Logiq E9 With Xdclear: Make Every Day ExtraordinaryDocument6 pagesLogiq E9 With Xdclear: Make Every Day ExtraordinarykritonNo ratings yet
- BCF Logiq S7 Sales SheetDocument2 pagesBCF Logiq S7 Sales SheetAnurak ThongbaiyaiNo ratings yet
- Strangulation Injuries - StatPearls - NCBI BookshelfDocument8 pagesStrangulation Injuries - StatPearls - NCBI BookshelfSteven CraigNo ratings yet
- Astm17 0303Document5 pagesAstm17 0303Edgardo Emilio CantillanoNo ratings yet
- Smartphone-based screening for diabetic retinopathy in IndiaDocument10 pagesSmartphone-based screening for diabetic retinopathy in IndiaVivi DeviyanaNo ratings yet
- 2010 Pegasus Lectures Seminar Order FormDocument1 page2010 Pegasus Lectures Seminar Order Formsigmoid123No ratings yet
- Lecture Notes by Saurabh Gupta SirDocument18 pagesLecture Notes by Saurabh Gupta SirlancetNo ratings yet
- Module 1Document58 pagesModule 1Weliam Jhay CoymeNo ratings yet
- Case 3 Care of Client With GI, PUD, Cancer, Liver FailureDocument33 pagesCase 3 Care of Client With GI, PUD, Cancer, Liver FailureNyeam NyeamNo ratings yet
- Curriculum Vitae: Name: Yousif Mohamed Yousif Abdallah Birth Date: Nationality: M. Status: Language: E. MailDocument17 pagesCurriculum Vitae: Name: Yousif Mohamed Yousif Abdallah Birth Date: Nationality: M. Status: Language: E. MailHassan NileNo ratings yet
- Principles of RadiologyDocument4 pagesPrinciples of RadiologyBenn BerriganNo ratings yet